 2001, Vol. 37
2001, Vol. 37文章信息
- 邓伯勋, 曾凤.
- Deng Boxun, Zeng Feng.
- 5个柑桔体细胞杂种的抗寒性研究
- STUDY ON FREEZING RESISTANCE OF 5 CITRUS SOMATIC HYBRIDS
- 林业科学, 2001, 37(6): 20-25.
- Scientia Silvae Sinicae, 2001, 37(6): 20-25.
-
文章历史
- 收稿日期:2000-10-09
-
作者相关文章


抗寒育种的研究是我国柑桔业中的主攻方向, 但过去主要依赖于体细胞突变, 即芽变和珠心苗变异。杂交育种工作受柑桔多胚性、童期长、遗传上的高度异质性, 以及柑桔大多性状为多基因控制的数量性状等因素的限制(邓秀新, 1991), 所以研究工作的进展非常缓慢。
20世纪70年代开始兴起的原生质体融合技术的成功为柑桔品种改良开辟了一条崭新的途径。自日本人F.Ohgawara等(1985)获得枳+“Trovita”甜橙体细胞杂种成功以来, 国内外柑桔育种学家们做了大量的柑桔体细胞杂交(亦称细胞融合或原生质体融合)工作。迄今为止, 在某桔育种方面已获得了100多个组合的体细胞杂种或胞质杂种。这些成功的组合中许多都含有粗柠檬、枳橙、酸橙、酸桔等具有不同抗性的砧木类型以及近缘属。其体细胞杂种作为砧木利用比作为接穗利用似乎更有价值, 尽早地鉴定这些体细胞杂种的抗性, 对于体细胞杂种的早日推广应用具有重要意义(甘霖等, 1995)。抗寒性作为柑桔栽培的北缘限制因子, 尤其需要尽早进行研究, 以便加速柑桔优良品种的推广与发展。
原生质膜与植物抗寒性关系密切, 1912年Maximov N.A指出低温伤害的关键是质膜。1932年Dexter ST, Totting harm WE对植物冻害后质膜的变化和电解质渗出现象进行了大量的研究, 从而建立了测定植物抗寒性的电导法。1940年Levitt J, Siminovittch D等的研究结果证明了细胞内外结冰均会破坏质膜。1981~1996年期间, 刘祖祺(1981)、简令成(1983)、王洪春(1989)、甘霖(1995)等研究结果均阐明了原生质膜的透性与植物的抗寒性成正相关。本研究用电导法研究细胞质膜在低温条件下的变化规律, 旨在探索几个原生质体融合再生植株的抗寒性的变异规律。
1 材料和方法 1.1 试验材料及设计试验的材料为华中农业大学柑桔基地中的5个体细胞杂种:伏令夏橙+宁波金柑(C.sinensis Osb.cv.Valencia+Fortunella crassifolia Swingle cv.Meiwa FV)、伏令夏橙+宜昌橙(C.sinensis Osb.cv.Valencia+ (C.ichangensis Swingle CV)、伏令夏橙+‘key’来檬(C.sinensis Osb.cv.Valencia+C.aurantifolia cv.Key lime KV)、哈姆林甜橙+飞龙枳(Citrus sinensis Osb.cv.Hamlin+Poncirus trifoliata Raf.cv.Flying Dragon HFd)、哈姆林甜橙+粗柠檬(C.sinensis Osb.cv.Hamlin+C.jamhiri L.HR), 以及这些组合的亲本, 即:哈姆林甜橙(HM)、粗柠檬(R1)、伏令夏橙(VI)、宁波金柑(FC)、宜昌橙(CI)、‘Key’来檬(key)。试材FV、HR、FC、RI、Key为1990, CV、KV、VI为1992, HM、CI、HFd为1995年的移栽苗, 生长势中庸。试材以单株为小区, 重复3次, 份精细管理和常规管理, 前者主要措施采取叶面喷肥、灌水和地面稻草覆盖, 其它同常规管理一样采取根际施肥, 病虫防治等。
1.2 材料处理试材采于1月中旬(12, 13, 14日) 9:00, 选用生长发育正常的春梢中部叶片。采后用流水冲洗15 min, 再用洗涤剂冲洗干净, 然后用蒸馏水洗2~3次, 最后用重蒸水洗1~2次; 将洗净的叶片用纱布擦干, 然后用直径0.30 cm打孔器将叶片打成小圆片, 把小圆片置于培养皿中加盖摆匀; 按每处理5个小圆片将上述圆片放入带塞10 mL刻度试管中, 每管中加入1滴重蒸水浸湿小圆片, 以保证叶片在处理过程中温度传递均匀且不受缺水胁迫。然后将试管盖好塞子, 对照置于室温下, 其余则按不同温度梯度放入冰瓶中。事先冰瓶内用碎冰调温, 从-5℃~-15℃设6个处理, 相邻两处理间温差均为2℃, 处理2 h后取出, 恢复0.5 h以解冻, 最后按1:5比例加入5 mL重蒸水。
1.3 测电导率用涡旋混合器将上述处理好的材料摇匀后, 在DDS-12A数字电导仪测电导率(Ci, Ti); 然后将试管置入高压灭菌锅内煮沸10 min, 冷却后再用涡旋混合器将材料摇匀, 测出电导率(Cf, Tf), 求出膜伤害率。
1.4 抗寒性测定 1.4.1 伤害率(injury %)
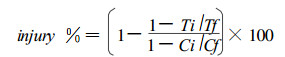
|
式中, Ci为煮前对照电导率; Cf为煮后对照电导率; Ti为煮前处理电导率; Tf为煮后处理电导率。
1.4.2 Logistic方程及拟合度的检验Logistic方程所描述的是呈“S”形变化规律的曲线。本研究温度对膜的伤害程度, 通过电导法测定, 观察到膜的透性随着温度的降低呈“S”型增大。所以用Logistic方程及拟合度进行检验, 莫惠栋(1983)、朱根海(1985)和苏维埃(1987)对此方法早就进行过讨论。
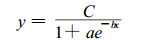
|
式中, y为伤害率; x为温度; c为伤害率渐近值(莫惠栋, 1983)。
所谓拟合度(ry′x), 即本实验中所得到的曲线与Logistic方程的符合程度, 这里用膜的伤害率与处理温度的相关系数进行判断。
1.4.3 拐点求法Logistic方程各参数求法均和朱根海等所做方法相同。
2 实验结果 2.1 低温对柑桔叶片细胞膜的影响随着抗逆研究的深入, 人们认为, 原生质膜与植物抗逆性关系最为密切。一般认为, 质膜稳定性愈强, 植物抗逆性也愈强(王洪春, 1985)。
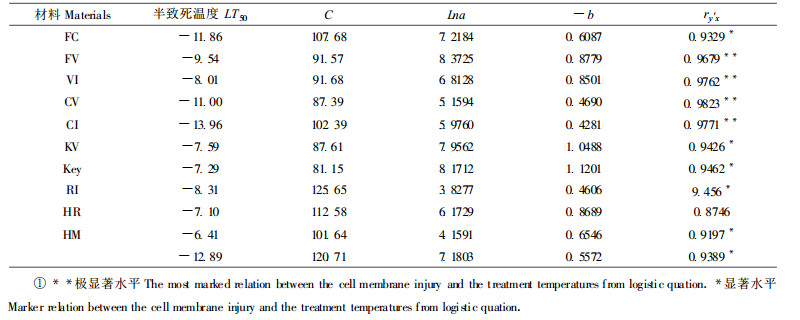
图 1~4所示曲线和表 2可见:随着温度的逐步降低, 柑桔体细胞杂种(KV、CV、FV、HR)细胞膜的伤害率变化规律均是开始变化小, 中间变化大, 最后又变化小。伤害率是反映膜稳定性的, 所以细胞膜在低温作用下变化的趋势也是呈现这种变化规律, 即这种变化呈较典型的“S”形曲线。研究中还表明了体细胞杂种和亲本的11种材料之间的半致死温度均有明显差异。即HFd为-12.89℃, CV为-11.00℃, KV为-7.5℃, HR为-7.10℃, CI为-13.96℃, FC为-11.86℃, FV为-9.54℃, RI为-8.31℃, VI为-8.01℃, Key为-7.29℃, HM为-6.41℃。尽管各个材料间其抗寒性表现不一, 但细胞膜受低温伤害的规律是基本一致的(图 1~4)。并且膜的伤害程度与柑桔的抗寒性呈负相关, 试材的抗寒性愈强, 细胞膜的伤害程度就愈轻。图 1~4中-11℃处理的结果表明:试材叶片细胞膜的伤害率依次是CI 24.41 < FC 43.51 < Key 82.04 < VI 86.69 < RI 01.38 < HM 111.28。利用方程、公式, 求出了不同柑桔试材的Logistic方程中的参数, 并对各处理温度与细胞膜伤害程度的关系用Logistic方程进行了检验(表 1), 其拟合度(ry′x)表明了FV、VI、CV和CI达极显著水平, HR表现不明显, 其余6个试材达显著水平。

|
图 1 低温对FV柑桔体细胞杂种和亲本叶片细胞膜的伤害程度 Fig. 1 Results of low temperatures for the cell membrane injury of citrus somatic hybrid(FV) and their parents leaves
|

|
图 2 低温对CV柑桔体细胞杂种和亲本叶片细胞膜的伤害程度 Fig. 2 Results of low temperatures for the cell membrane injury of citrus somatic hybrid(CV) and their parents leaves
|

|
图 3 低温对KV柑桔体细胞杂种和亲本叶片细胞膜的伤害程度 Fig. 3 Results of low temperatures for the cell membrane injury of citrus somatic hybrid(KV) and their parents leaves
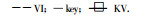
|

|
图 4 低温对HR柑桔体细胞杂种和亲本叶片细胞膜的伤害程度 Fig. 4 Results of low temperatures for the cell membrane injury of citrus somatic hybrid (HR) and their parents leaves

|
|
|
|
|
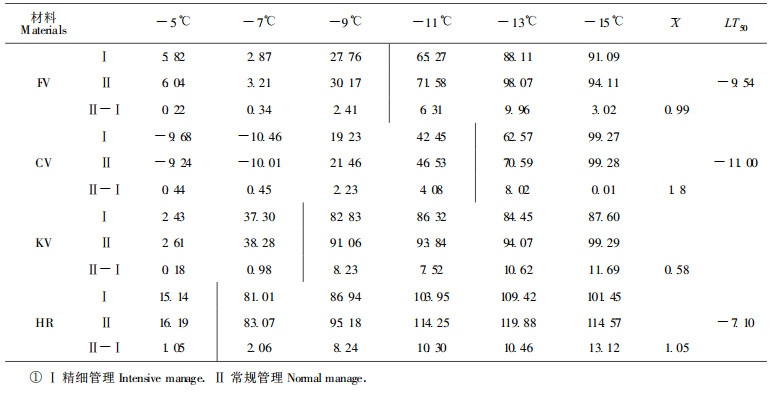
表 1表明, 4个柑桔体细胞杂种的抗寒性或半致死温度(LT50)介于其两个亲本之间, 并且较半致死温度较高的亲本一方有所降低, 如宁波金柑(FC)的LT50为-11.86℃, 伏令夏橙(VI)的LT50为-8.01℃, 而宁波金柑+伏令夏橙(FV)的LT50为-9.54℃。显而易见, FV柑桔体细胞杂种的抗寒性介于两亲本之间, 较抗寒性弱的亲本(VI)提高抗寒能力1.53℃, 较抗寒性强的亲本FC的抗寒性降低2.32℃。CI与VI组合杂种(CV)的抗寒性同样介于两亲本之间, 抗寒力为-11℃, 较抗寒性弱的亲本(VI)提高抗寒能力3℃, 但较CI亲本降低了2.96℃。VI和Key组合杂种(KV)的抗寒性为-7.59℃, 较抗寒性弱的亲本(Key)提高抗寒能力0.3℃, 比亲本(VI)降低抗寒力0.42℃。HM和RI组合杂种(HR)的抗寒力(-7.12℃)也较抗寒力低者(HM)提高0.69℃, 较另一亲本(R1)的抗寒力降低0.21℃。体细胞杂种(HFd)在-5℃, -7℃, -9℃, -11℃, -13℃的6种低温处理中, 其细胞膜的伤害率依次为0.83 (±2.26), 13.15 (±2.25), 15.37 (±3.91), 46.15 (±7.73), 55.47 (±6.50), 91.50 (±8.89), 结果表明了半致死温度为-12.89℃, 而亲本HM的半致死温度则为-6.41℃ (图 4)。飞龙枳是国外用的砧木, 其特点是极矮化, 抗寒性强, 本试验无材料分析。所以结果只能表明HFd的抗寒性较亲本(HM)明显地提高6.48℃, 并且较其它砧木矮化, 这是几个体细胞融合组合中提高抗寒力极为显著的杂种。总之, 体细胞杂种的抗寒性取决于两亲本的抗寒能力, 较抗寒性弱者的亲本有所提高, 低于抗寒性强者亲本。并且4组体细胞杂种组合均是如此规律。图 1~4中4个组合的Logistic方程所示曲线表明了上述几个体细胞杂种抗寒性介于两亲本之间的趋势, 同时也显现了细胞膜在不同温度条件下的伤害程度及变化规律, 杂种与其亲本基本一致。
从半致死温度来看(表 1), 哈姆林甜橙+飞龙枳的体细胞杂种(HFd)为-12.89℃, 宜昌橙+伏令夏橙的体细胞杂种(CV)为-11.00℃, 伏令夏橙+宁波金柑的体细胞杂种(FV)为-9.54℃, ‘Key’来檬+伏令夏橙的体细胞杂种(KV)为-7.59℃, HR体细胞杂种为-7.10。由此可见, HFd以及CV两个体细胞杂种的抗寒性较其它组合强。
2.3 柑桔体细胞杂种抗寒性与栽培管理的关系本研究探讨了柑桔体细胞杂种的抗寒性是否受管理水平的影响。表 2中, 4个柑桔体细胞杂种在精细管理(Ⅰ)和常规管理(Ⅱ)条件下, 细胞膜对低温的忍受程度均无明显差异。在半致死温度内, Ⅰ处理比Ⅱ处理的细胞膜伤害率仅少1.1%左右, 其差异分别表现为FV 0.99%, CV 1.8%, KV 0.58%, HR 1.05%。两种处理对细胞膜伤害程度的差异虽然很小, 但4个体细胞杂种在精细管理条件下似乎有可能提高抗寒性的现象。
3 讨论植物的细胞膜是细胞进行物质运输、能量交换、信息辩别与信息传递的重要途径, 它对生命活动起着极为重要的作用。所以研究体细胞杂种细胞膜与抗寒性的关系, 是认识体细胞杂种抗寒机理的重要内容。尽管细胞膜的抗寒功能依赖于膜的流动性(苏维埃, 1983), 以及与细胞膜的组成成分, 如膜蛋白、膜磷脂、脂脂肪酸和酸系统等结构物质均有关。但细胞膜功能的正常发挥与低温的强度关系十分密切(Mccort, 1987;Mannan, 1987)。强低温的作用可以破坏细胞膜的结构, 使其膜的透性加大, 细胞内的K+和葡萄糖等物质外渗, 使细胞膜的功能丧失; 最终影响植物体的生存。膜受害程度轻则透性的变化可逆, 重则不能恢复正常。这种变化明显地出现在外部形态之先。所以用电导法测定低温处理后的柑桔体细胞杂种的叶片细胞内电解质的渗出量, 是研究其抗寒性的主要生理指标。
从本实验中4个柑桔细胞杂种及亲本的细胞膜透性与杂种抗寒性的关系, 可以表明, 抗寒性强的亲本的细胞膜受低温伤害的程度轻, 抗寒性弱的亲本则反之。杂种抗寒性介于两亲本之间, 比抗寒性强的亲本低, 较抗寒性弱的亲本强, 并且4个杂种的抗寒性与亲本组合的抗赛性呈正相关。结果还表明了体细胞融合抗寒性育种似乎没有杂种优势的趋势显现, 仅强于弱者的抗寒性, 偏向抗寒性强的亲本一方。可见柑桔体细胞融合杂种的抗寒力取决于亲本的抗寒性及遗传力。此外, 本实验结果还表明了杂种的抗寒能力几乎不受栽培管理水平的影响, 但精细管理似乎有可能提高杂种抗寒性的现象存在。
5个柑桔体细胞杂种抗寒性的研究结果表明, HFd抗-12.89℃; CV抗-11.00℃; FV抗-9.54℃; KV抗-7.59℃; HR抗-7.10℃。体细胞杂交是一无性过程, 后代不存在基因分离现象, 亲本中由显性基因或共显性基因控制的性状在体细胞杂种中有可能得到表现, 而柑桔的抗寒性是由显性基因控制的性状(甘霖, 1995)。因此, 通过体细胞杂交的再生植株的抗寒性状是稳定的。
邓秀新. 1991. 柑桔原生质体融合及遗传转化的研究进展. 果树科学, 8(1): 51-58. |
甘霖, 孙中海, 邓秀新, 等. 1995. 柑桔体细胞杂种的抗性研究. 园艺学报, 22(3): 209-214. DOI:10.3321/j.issn:0513-353X.1995.03.017 |
郭金铨. 1978. 在冻害过程中咖啡离体叶细胞透性变化的研究. 植物生理学报, 53: 199-203. |
简令成. 1983. 生物膜与植物寒害和抗寒性的关系. 植物学通报, 1: 17. |
刘祖祺. 1981. 电导法鉴定柑桔耐寒性的试验. 南京农业大学学报, 2(6): 32. |
莫惠栋. 1983. Logistic方程及其应用. 江苏农学院学报, 4(2): 53-57. |
苏维埃. 1983. 膜脂不饱和度在水稻抗冷性中的作用. 科学通报, 28(3): 373-376. |
王洪春. 1985. 植物抗逆性与生物膜结构功能研究的进展. 植物生理学通讯, 1: 60-66. |
王洪春. 1989. 植物耐寒性及防寒技术. 北京: 学术书刊出版社, 114-117.
|
朱根海, 刘祖祺, 朱培仁. 1986. 应用Logistic方程确定植物组织半致死温度的研究. 南京农业大学学报, (3): 11-15. |
Dexter ST, Totting harm WF. 1932. Investigations of the hardiness of phants by measurement of electrical conductivity. Plant physiol, 7: 63. DOI:10.1104/pp.7.1.63 |
Kechchic D O. Methods of determing cold hardiness and cold injury in citrus. Proceedings of the first international citrus symposium. 1968, Vol, 2: 559~563
|
Levitt J, Siminovitch D. 1940. The relation between frost resistance and the physical state of protoplasm. Can. J. Res. C., 18: 550. |
Lyons J M. 1973. Chilling injury in plants. Ann. Rev. Plant physioll, 24: 445-445. DOI:10.1146/annurev.pp.24.060173.002305 |
Mannan R M. Changes in the cationtic regulation of structure and function in thylakoids isolated from wheat seedlings treated with BASF13-338 plant physiol, 1987, 85: 17~19
|
Mccourt P. 1987. The effects of rejuicedam ounts of lipid unsaturation on chloroplast ultrastructure and photosynthesis in a mutant of Arabidopsis. Plant physiol, 84: 353-360. DOI:10.1104/pp.84.2.353 |