 2001, Vol. 37
2001, Vol. 37文章信息
- 陈存及, 邱尔发, 梁一池, 范辉华, 陈东海.
- Chen Cunji, Qiu Erfa, Liang Yichi, Fan Huihua, Chen Donghai.
- 毛竹不同种源光合特性研究
- STUDY ON THE PHOTOSYNTHETIC CHARACTERS OF PHYLLOSTACHY HETEROCYCLA CV.PUBESCENS PROVENANCES
- 林业科学, 2001, 37(6): 15-19.
- Scientia Silvae Sinicae, 2001, 37(6): 15-19.
-
文章历史
- 收稿日期:2000-06-07
-
作者相关文章


2. 福建省建瓯市南雅林业站 建瓯 353100
2. Jian′ou Nanya Forest Station, Fujian Province Jian′ou 353002
毛竹(Phyllostachys heterocycla cv. pubescens)起源于我国, 是我国南方最主要的经济竹种。在竹类遗传改良方面, 由于竹子本身的克隆特性, 研究不多。20世纪70年代, 张光楚等进行了竹类杂交育种和染色体等方面的研究(张光楚等, 1986;广东省林科所营林室育种组, 1979)。美国Georgia大学Friar.E用RFlPS方法研究刚竹属中20竹种(包括毛竹)的种内及种间遗传变异和演化(Friar, 1994), 有台湾学者从DNA多态现象研究台湾毛竹遗传因子的变化, 采用RAPD分析法对来自全岛屿的176个样品家系进行了分析鉴定, 分别用BRAPD, 1小卫星引物和3大卫星引物放大DNA基因片断, 并进行了聚类分析和联合分析, 结果把这176个家系样品归为9类, 样品中遗传基因变化小(Lai et al., 1994)。在竹类种源研究方面福建林学院首先开展了毛竹种源试验(陈存及等, 1999;邱尔发等, 1999), 而竹类光合特性的研究也很少(黄启民等, 1989)。毛竹不同种源的光合作用是否存在变异, 是毛竹研究领域中一个重大问题。
种源选择能有效地利用种源的优良性状(马德华等, 1982;刘来福等, 1984)。各地的树种、种源经过长期的自然选择, 可能在某些性状方面存在遗传变异。俞新妥教授于1956年在福建南平开展马尾松种源试验, 20年生时, 广西种源比本地种源树高大14.5 %, 材积大44.2 %(沈熙环, 1992;俞新妥等, 1985)。这说明种源间差异明显, 不同的种源在各地的性状表现不同, 种源间的遗传差异常常是个体间遗传差异的数倍。所以, 数十年来, 种源试验被看作是树种改良的基础研究予以优先考虑和实施。从竹类的自然变异看, 竹林的产量、质量、笋的产量、品味、有效成分的含量、加工性能以及出笋期等特性都存在着明显的差异。所以, 通过毛竹种源试验, 分析不同种源的光合特性, 探索不同产地毛竹群体的变异模式, 选择品质优良的高光效毛竹种源, 是一项非常有意义的工作。
1 试验地及试验材料 1.1 试验地自然条件试验地设于建瓯市南雅镇头村林场, 地理位置为东经118° 05′, 北纬26° 44′。建瓯市年平均气温19.4 ℃, 年平均降水量1726.6 mm, 1月份平均气温9 ℃, 7月份平均气温28.5 ℃, 绝对最高气温44 ℃, 最低气温-6 ℃, 全年无霜期270 d。试验林所在山场海拔150 ~ 200 m, 平均坡度25°, 土壤为山地红壤, 土层厚度大于100 cm, 表土层15 ~ 20 cm, 立地质量综合评价为Ⅱ级。
1.2 试验材料根据我国毛竹分布区域划分为3个种源带:高于北纬30°为北带、低于北纬25°为南带、北纬25°~ 30°之间为中带。共收集分布在3个种源带的8省16个毛竹种源母竹作为试验材料(表 1)。母竹的选择条件包括毛竹年龄、胸径、枝下高、留枝盘数、保留来鞭、去鞭长度等指标严格控制基本一致。母竹挖掘后宿土包扎, 并立即运输到试验地栽种。
|
|
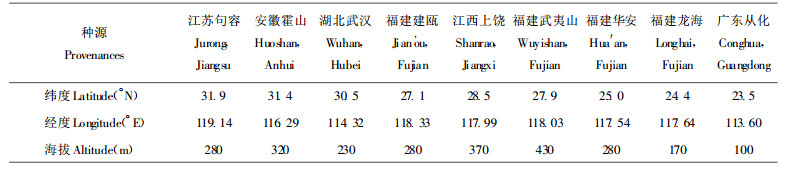
从16个种源试验林中, 每一个种源带各选择3个种源, 共9个种源进行光合作用测定。9个种源地理位置见表 1。
2 研究方法参见文献(北京师范大学生物大学教研室, 1979;北京大学生物系化学教研室, 1979)。
2.1 测定时间分别于1996年5月下旬(21 ~ 29日)、7月中旬(14 ~ 20日)、10月上旬(6 ~ 12日)、11月中旬(22 ~ 29日)、12月下旬(23 ~ 29日)测定; 分析样品选取1996年新竹中部向阳面竹叶。
2.2 净光合速率和暗呼吸速率测定采用北京北分麦哈克分析仪器有限公司生产的GXH-305便携式CO2分析仪测定离体枝条的净光合速率; 用黑布遮光测定暗呼吸速率。
2.3 光补偿点测定测定0-10000Lux光强下的光合强度, 测定扣除叶室吸收后到达叶面的实际光强, 以实际接受的光强计算离体枝条的光补偿点。
2.4 叶面积测定采用美国产的Li-3000叶面积仪测定叶面积(Li-cor, 1981; Ludwing, 1965)。
2.5 光合作用条件控制光强:用碘钨灯为光源, 交流电子稳压器稳定电源, 照度计测光强。
温度:用空调调节室内温度, 叶室上方用隔热水槽降温, 用温度计测定温度。
湿度:在叶室中用湿棉花调节湿度。
气源:采集离地面5 m的稳定空气。
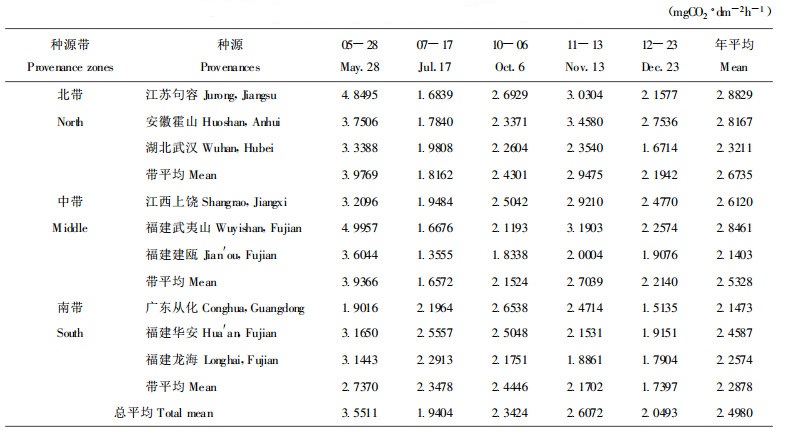
3 结果与分析 3.1 净光合速率年周期变化净光合速率是反映光合能力的重要指标, 从各种源来看, 新竹光合作用在一年中有两个峰值, 第一次都是在5月, 新竹形成, 展叶完成时。但第二次峰值出现的时间有所差异, 总体上, 南带3个种源第二次净光合速率峰值出现较早, 都在10月份, 其它种源相对较迟。这可能是由于南带种源对低温较为敏感, 光合能力受低温制约较大, 当低温来临时, 可能使酶活性下降, 从而导致净光合速率较早出现下降。从净光合速率年平均值看, 变化范围为2.1403 ~ 2.8829 mg CO2·dm -2h-1, 变化幅度不大, 最大的为江苏句容种源, 最小的为福建建瓯种源。从种源带的变化看, 虽然各种源在一年中具有不同变化, 但总体上, 净光合速率有随纬度的升高而增高的趋势, 即北带>中带>南带。
|
|
|
|
方差分析结果表明, 各种源及种源带之间净光合速率差异不显著(F =1.64 < F0.05(8.32)=2.24), 但5月份与7月、10月、11月、12月份净光合速率均达极显著差异水平(F =13.46 >F0.01(4, 32)= 3.97)。这说明不同季节的净光合速率不同, 而不同种源之间差异不大。这可能与不同季节酶活性有关, 5月份展叶期光合酶活性较高。
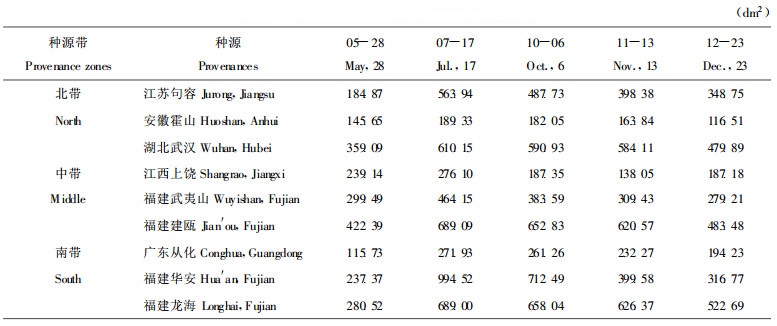
3.2 单株光合生产率单株光合生产率为单株叶面积与净光合速率的乘积。从表 4可见, 单株光合生产率在7 ~ 10月份最高, 其中以福建华安种源为最大, 福建龙海、湖北武汉和福建建瓯等种源次之, 其净光合速率都大于月平均值, 最小的为安徽霍山种源。
|
|

经方差分析, 种源间单株光合生产率达极显著差异(F =5.57 >F0.01(8, 32)=3.13, F =13.80 >F0.01 (4, 32)=3.97), 但不同月份间差异不显著(F =2.54 >F0.05(4, 32)=2.67), 这主要是由于不同种源单株叶面积不同所致。经进一步多重比较, 福建华安种源与江西上饶、广东从化和安徽霍山种源, 福建建瓯种源与广东从化、安徽霍山种源, 福建龙海种源与广东从化、安徽霍山种源, 湖北武汉种源与广东从化、安徽霍山种源均达显著差异水平。
3.3 不同种源新竹光补偿点比较光补偿点反映不同种源毛竹利用弱光的能力。从图 1可见9个种源光补偿点的变幅在570 ~ 913 Lux, 光补偿点最低的为江西上饶种源, 只有各种源平均值的84.2 %, 其次为武夷山、福建华安、福建建瓯、福建龙海、广东从化等种源, 分别只有平均值的0.89、0.91、0.95、0.96和0.97倍, 最高的为安徽霍山种源, 是各种源平均值的1.35倍。从种源带看, 北带种源光补偿点明显高于其它种源, 以中带种源为最低, 平均为603 Lux。这说明中带种源具有较强的利用弱光能力, 它在一天中能更有效的利用光能合成碳水化合物, 而北带种源则相反。

|
图 1 不同种源光补偿点 Fig. 1 The light compensation point of different provenances 1江苏句容Jurong, Jiangsu 2安徽霍山Huoshan, Anhui 3湖北武汉Wuhan, Hubei 4江西上饶Shangrao, Jiangxi 5福建武夷山Wuyishan, Fujian 6福建建瓯Jian′ou, Fujian 7广东从化Conghua, Guangdong 8福建华安Hua′an, Fujian 9福建龙海Longhai, Fujian 10平均值Mean |
新竹光合作用在一年中有两个峰值, 第一次所有种源都是在5月, 但第二次峰值南带种源出现较早, 都在10月份, 其它种源相对较迟。净光合速率年平均值变化范围为2.1403 ~ 2.8829 mgCO2·dm -2h-1, 最大的为江苏句容种源, 最小的为福建建瓯种源。各种源带净光合速率有随纬度的增升高而增大的趋势。
各种源单株光合生产率在7至10月份最高, 其中以福建华安种源为最大, 福建龙海、湖北武汉和福建建瓯等种源次之, 其净光合速率都大于月平均值, 最小的为安徽霍山种源。
各种源光补偿点的变幅在570 ~ 913 Lux, 其中最低的为江西上饶种源, 只有各种源平均值的84.2 %, 最高的为安徽霍山种源, 是各种源平均值的1.35倍。中带种源能更有效的利用光能, 光补偿点的平均为603 Lux, 而北带种源光补偿点明显高于其它种源。
北京大学生物系化学教研室编.生物化学实验指导.北京: 人民教育出版社, 1979
|
北京师范大学生物系植物教研室组主编.植物生理学实验指导.北京: 人民教育出版社, 1980, 11
|
陈存及, 邱发尔, 等. 1999. 毛竹种源春笋营养成份分析研究. 竹子研究汇刊, 18(1): 6-11. |
广东省林科所营林室育种组. 1979. 竹子有性杂种的初步选择. 林业科技通讯, (1). |
黄启民, 等. 1989. 不同条件下毛竹光合作用的研究. 竹类研究, 2: 8-17. |
梁一池. 1997. 树木育种原理与方法. 厦门: 厦门大学出版社.
|
刘来福编著.作物数量遗传.北京: 农业出版社, 1984, 149~162, 206~284
|
马德华编著.植物育种的数量遗传学基础.南京: 江苏科学技术出版社, 1982, 245~279
|
邱尔发, 陈存及, 等. 1999. 竹种源叶片性状年动态研究. 福建林学院学报, 19(1): 87-90. DOI:10.3969/j.issn.1001-389X.1999.01.025 |
沈熙环编著.林木育种学.北京: 中国林业出版社, 1992
|
俞新妥, 等. 1985. 杉木种源地理位置的数学模型. 福建林学院学报, 5(2): 1-5. |
张光楚, 陈富枢. 1986. 竹类杂交育种的研究. 竹类研究, 5(3): 48-53. |
Friar E, Kochert G(a).A study of genetic variation and evolution of Phyllostachys (Bambusoideae; Poaceae)using nuclear restriction fragment length polymorphisms.Theoretical and Applied Genetics 1994, 89(2~3): 265~270
|
Lai c c, Hsiao J Y. 1994. Genetic variation of Phyllostachys pubescens(Bambusoidege, Poaceae)in Taiwan based on DNA polyorphisms. Botanical Bulletin of Academia Sinica, 38(3): 45-152. |
LI-COR.Inc, Instruction manual of Li~3000 portable area meter, 1981
|
Ludwing L J et al..Photosynthesis in artificial communities of cotton plants in relation communities of cotton plants in relation to leaf area.J.Biol, Sci.,
|