 2001, Vol. 37
2001, Vol. 37文章信息
- 王朝晖, 费本华, 任海青, 郝刚.
- Wang Zhaohui, Fei Benhua, Ren Haiqing, Hao Gang.
- 长江滩地立木腐朽杨树与正常杨树生长与材性的比较研究
- COMPARATIVE RESEARCH ON GROWTH AND WOOD PROPERTIES BETWEEN ROTTEN POPLAR TREE AND NORMAL ONE ON FLOOD BEACHES ALONG YANGTZER RIVER
- 林业科学, 2001, 37(5): 113-119.
- Scientia Silvae Sinicae, 2001, 37(5): 113-119.
-
文章历史
- 收稿日期:2001-01-09
-
作者相关文章


2. 安徽省阜阳市颖州区农林局 阜阳 236000
2. Fuyang Agriculture and Forest Bureau, Anhui province Fuyang 236000
长江中下游地区位于亚热带季风气候区, 生物、气候、土壤资源丰富, 是我国粮、棉、油的重要产区。然而, 近年来由于长江上游地区森林生态系统被破坏, 造成了大量的水土流失, 恶化了滩地的生态环境, 为吸血虫的中间宿主———钉螺提供了良好的孽生环境, 造成血吸虫病的流行。随着“以林为主, 灭螺防病, 综合治理和开发滩地”这一科学的生态工程(江泽慧等, 1995; 彭镇华等, 1995)的实施, 不仅减少了血吸虫病疫性、控制了水土流失, 还增加了森林资源, 提高滩地的利用价值。杨树是一个世界性树种, 黑杨派在杨树栽培中有着重要的位置, 是世界中纬度地区栽培面积最大的树种之一, 它适宜于在平原地区、河漫滩地、围垦区等废闲土地上进行成片造林, 投入少, 见效快, 充分利用了地力, 经济效益十分显著。杨树作为重要的工业用材树种之一, 在胶合板工业、造纸工业等方面已得到广泛的应用。但是杨树作为滩地主要造林树种之一, 经常发生立木腐朽现象, 大大降低了杨树木材利用价值和农民的造林积极性。为了更好的开发、推广杨树在滩地上的合理利用, 本文对长江滩地立木腐朽杨树与正常杨树生长和材性进行比较研究, 探明菌丝在杨树木材中的侵入和分布情况, 从而为进一步消除或减少杨树腐朽提供基础。通过对腐朽材和正常材结构和性质差异的研究, 为更好的利用杨树木材提供科学的依据。
1 材料与方法 1.1 材料 1.1.1 试材的采集首先对“长江中下游滩地综合治理和开发”课题固定实验点安徽省安庆市红星乡林场意杨林进行踏查选点, 由于地势高低不同, 根据去年水淹的最高水位作为标准, 选取地势高低不同的3个标准地A1、A2、A3, 其中A1的地势最低, A3最高, A2居中。查数其每个标准地内的腐朽木株数。结果如表 1。
|
|

从表 1中可看出, 地势越低, 腐朽木的株数越多, 这主要是由于地势低, 受水淹的时间长, 积水时间长, 加速了立木杨树树干腐朽。A1内腐朽木最多, 实测其全部树木的树高、胸径, 并给每棵树标号、记录其腐朽木号。把正常木与腐朽木分开, 根据其胸径的大小分成大、中、小3级, 计算各级树木的平均胸径, 以此作为选取标准木的主要标准, 其结果如表 2。
|
|
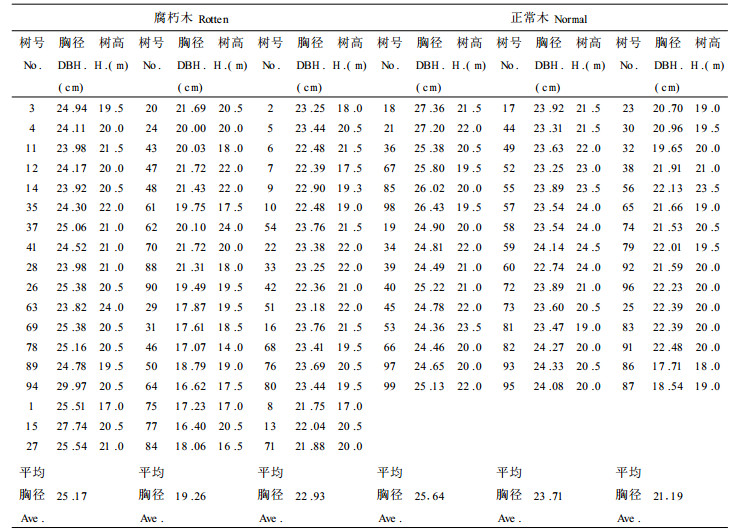
根据各级平均胸径, 且以树高和干形作为参照选取9号、30号、50号、67号、72号、78号为标准木。通过对腐朽木和正常木伐倒调查, 腐朽木一般从树干基部开始, 一直蔓延到胸径以上部位, 基部腐朽最严重, 越到上部越轻微, 同时腐朽木树干均不保持直立, 而是朝某一方向倾斜, 树干基部腐朽的杨树一般没有根腐现象。
1.1.2 试材的制作将圆盘刨平以便查数其年轮, 求其晚材率及胸径生长曲线。沿南北向截取通心试条1根, 取下再截成4部分, 第1部分留存, 第2部分根据标准将其加工成20 cm×20 cm×20 cm的试样, 用于测含水率、干缩率、湿胀率、基本密度。
1.2 方法 1.2.1 生材含水率的测定样木伐前在树干上标定南北线。伐倒后, 在每株树的基部、1.3 m、2 m处分别截取厚度为30 cm的圆盘, 并给每个圆盘标号。其中基部的向上截取, 1.3 m、2 m处分别向下截取。1.3 m处圆盘锯下后, 立即在树干的内部和外部分别截取木块2块, 称重后妥善包装带回测含水率。
1.2.2 木材含水率的测定方法按国家标准GB 1931-91 《木材含水率测定方法》的规定, 将加工好的试样(清除干净附在试样上的木屑、碎片等)称重, 然后放入烘箱内, 在103±2 ℃的温度下烘8 h后, 从中选定2~3个试样进行第1次试称, 以后每隔2 h试称1次, 至最后2次称量之差不超过0.002 g时, 即从烘箱中取出, 放入装有干燥剂的玻璃干燥器内, 冷却至室温后取出称重。
1.2.3 木材年轮宽度的测定方法按国家标准GB 1930-91 《木材年轮宽度和晚材率测定方法》的规定, 在试样端面上, 按径向划一直线, 沿直线测出整年轮部分的总宽度, 准确至0.01 mm。并数出测量范围内的整年轮数。
1.2.4 木材基本密度的测定GB 1933-91 《木材密度测定方法》在胸高圆盘上按1~3轮, 4~6轮, 7~9轮等依次取样, 试样与测定木材含水率相同, 腐朽杨树取样试块为完整立方体, 略带腐朽, 用排水法测定, 把3株正常木与3株腐朽木的平均值做为各年轮的基本密度值。
1.2.5 木材干缩率的测定按照GB 1932-91 《木材干缩性测定方法》的规定。
1.2.6 木材显微结构的分析用常规染色切片法制作永久切片在光学显微镜下观测并拍照; 另外先制作50~70μm切片, 然后用真空喷镀仪喷金, 在Phillip扫描电镜下观测。
2 结果与讨论 2.1 腐朽对树木生长的影响树木的生长包括高生长和径生长, 它们有着自身的特性, 同时又受外界条件的影响。根据标准地A1的腐朽木与正常木树高和胸径, 进行方差分析, 结果如表 3:
|
|
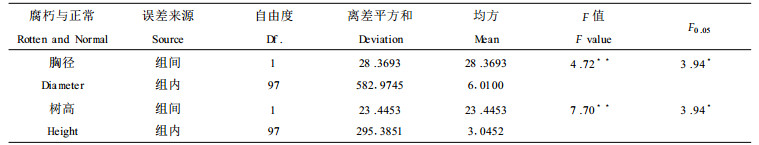
经单因素方差分析表明, 在0.05水平上, 腐朽材与正常材在树高、胸径上差异显著。即腐朽对杨树的高生长和胸径生长都有一定的影响, 从F值大小可见, 腐朽对高生长影响大于腐朽对胸径生长的影响。
2.2 腐朽木与正常木年轮宽度的比较年轮宽度和晚材度是木材宏观构造上的两个重要指标, 与木材密度存在紧密的相关关系, 通过对生长轮的分析, 可以预测许多指标。腐朽木与正常木年轮宽度的径向变异如图 1所示:

|
图 1 年轮宽度的径向变异 Fig. 1 Radial variation of the width of yearly ring
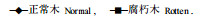
|
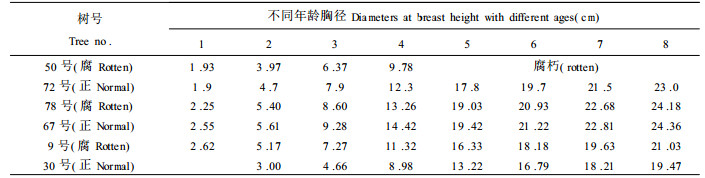
图 1表明, 生长轮在不同的年龄阶段有不同的年轮宽度, 正常木与腐朽木的年轮生长趋势基本一致(第4a除外)。由于第1a的年轮宽度很小, 因此在图中没有第1a的年轮宽度。图中从第2a开始年轮宽度逐年上升, 到第6a后开始下降。这个结果与鲁西杨树的生长规律和胸径粗生长过程的研究结果(曾其蕴等, 1990; 刘寿坡等, 1992)相一致。这种生长轮宽度的变化规律可能是意杨自身生长特性决定的。同时, 这也说明腐朽只对树木的生长势产生影响, 并不影响它的生长规律。根据杨树的这一生长规律, 对表 5采用y=a +b/x和y=a·e-b/x两种方程对其径向生长进行模拟, 结果如下表:
|
|
|
|
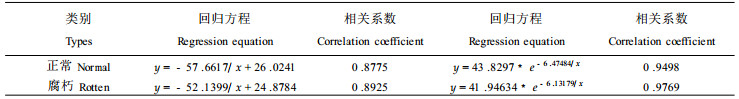
对两种回归方程进行比较, 两种回归方程的相关系数表明, 无论是腐朽木的还是正常木, 都是采用y=a·e-b/x对杨树的径向生长描述比较好。对于腐朽木, 可采用y=41.94634·e-6.13179x, 对于正常的可采用y=43.8297·e-6.47484x。分析与讨论:(1)此方程只适用于长江中下游滩地; (2)由于只采用前9 a的数据, 故只可用于预测杨树的早期胸径生长。
2.3 腐朽木与正常木含水率的比较木材的含水率包括生材含水率和气干含水率, 湿材含水率, 其中生材含水率对于木材运输和贮存等具有一定实际意义。生材中的水分主要是自由水和吸着水, 化合水的数量极少, 可忽略不计。对于不同的树种其生材所含吸着水数量接近, 所以生材含水率的高低主要是自由水含量不同的结果。表 6为样木要伐倒时所采样的含水率表。
|
|
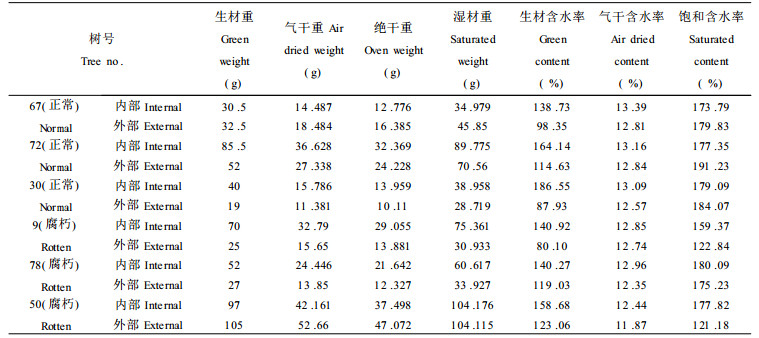
从表 6、表 7中可看出正常木的内部生材、气干材含水率大于外部, 而内部位饱和含水率略低于外部, 这是由于正常木内部所含树胶比外部多, 且树胶覆盖于管间纹孔、导管-木射线纹孔之上, 使纹孔被堵塞, 使木材吸水受阻。对于腐朽杨树, 内部生材、气干含水率大于外部, 这与正常木一致, 但腐朽材内部位饱和含水率大于外部, 这与正常材恰相反, 这主要是由于腐朽材外部的吸水性降低而使其饱和含水率大大降低所致。同时腐朽杨树内部平均生材含水率低于正常杨树内部平均生材含水率, 而腐朽杨树外部平均生材含水率略高于正常杨树外部平均生材含水率, 这使得腐朽杨树内部与外部平均生材含水率之差远低于正常杨树这可能是真菌侵蚀造成杨树纤维细胞、木细胞被破坏细胞腔被子粘液和微粒所填充, 致使水份进入较少, 内部木材输送水分功能减弱所致。而图中表明, 气干材含水率的变化趋势与基本密度相一致。
|
|

木材密度是判断木材各项力学强度的重要指标。对于正常木材在含水率相同的情况下, 密度愈大的木材强度也愈大, 木材强度与木材密度关系密切。另外全干材的空隙度随着全干密度的增大而减小, 木材的最大吸水率也随着全干密度的增大而减小。因此, 了解腐朽对木材密度的影响很有必要。把正常木与腐朽木的密度进行平均, 结果如图 2:

|
图 2 正常木和腐朽木基本密度径向变异 Fig. 2 Radial variation of average basic density comparing rotten wood with normal
|
从图 2中可看出, 杨树基本密度径向变异的趋势是自髓心到树皮先增加后减小, 这与研究杨树林分内木材比重径向变异四种模式的结果(Loo et al., 1985)相一致。
一般来说完全或部分腐朽的木材密度低于正常木材。但上图表明, 立木腐朽杨树木材的密度比正常材略高, 这主要是由于其密度试样取自腐朽杨树中基本完好的木材(略带腐朽)。利用显微切片观察进一步研究其原因, 从图版Ⅰ中图 6及图版Ⅱ中图 15、16来看, 立木腐朽杨树木材横切面中出现了带状胞壁加厚, 同时腐朽菌的菌丝周围均匀包绕着一层粘液状外层覆盖物, 这层覆盖物充满整个细胞腔, 其中还有许多微小颗粒, 因而腐朽杨树木材中基本完好的部分由于产生异常胞壁加厚, 同时从图版Ⅰ中图 2、5及图版Ⅱ中图 1、4的比较可见, 腐朽材导管比量比正常材低, 由于这两个原因使得其密度比正常木高, 但不能据此说明腐朽木的强度高于正常木。从整体而言立木腐朽杨树木材的强度仍低于正常木。

|
图版Ⅰ Plate Ⅰ 1.正常木基部圆盘; 2.正常木横切面(×63);3.正常木横切面(×400);4.腐朽木基部圆盘; 5.腐朽木横切面应拉木区域(×63);6.腐朽木横切面应拉木区域(×400);7.腐朽木横切面腐朽区域(×40);8.腐朽木横切面腐朽区域(×400)。 1.Normal poplar round sample; 2.Normal poplar cross section (×63);3.Normal poplar cross section(×400);4.Rotten poplar round sample; 5. Cross section of reaction(×63);6.Cross section of reaction(×400);7.Cross section of rotten(×40);8.Cross section of rotten(×400) |

|
图版Ⅱ Plate Ⅱ 1.正常木电镜横切面(×50);2、5.正常木早材电镜横切面(×1010、×5000);3、6.正常木晚材电镜横切面(×1010、×5000);4.腐朽木电镜横切面(×41);7、8.腐朽木电镜横切面应拉木区域(×1010、×2500);9.腐朽木电镜横切面菌丝(×1010);10.腐朽木电镜径面导管内菌丝(×406);11.腐朽木电镜径面射线内菌丝(×810);12.腐朽木电镜弦面纤维内菌丝(×810)。 1.Normal poplar cross section(×50);2、5.Cross section of earlywood(×1010、×5000);3、6.Cross section of latewood(×1010、×5000);4.Rotten poplar cross section(×41);7、8.Cross section of reaction(×1010、×2500);9.Mycelium in cross section(×1010);10.Mycelium in vessel(×406);11.Mycelium in ray cell (×810);12.Mycelium in fiber cell(×810). |
木材的干缩影响到木材的加工利用, 根据干缩率的大小可以决定制材的加工余量, 差异干缩是弦向干缩系数与径向干缩系数的比值, 其数值反映了木材弦向干缩与径向干缩的差别程度。差异干缩愈大, 木材愈容易开裂和变形; 差异干缩愈小, 木材的尺寸和体积稳定性愈好。腐朽木与正常木的干缩差异性分析如表 8:
|
|
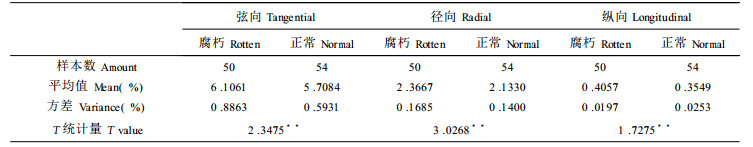
表 8表明, 腐朽木与正常木在弦向、径向、纵向上都存在显著性差异。且干缩率都比正常材高, 其中弦向高6.97%, 径向高0.96%, 纵向高14.31%。这主要是由于腐朽木都为应拉木, 其胶质纤维数比正常材高。另外, 不论是腐朽木还是正常木, 其纵向干缩率都很小, 而横向干缩较大。其中弦向干缩是径向干缩的两到三倍之间, 是纵向干缩的十倍左右, 这与前人的研究结果相一致。腐朽木与正常木各向的干缩率变化趋势一致, 都为自髓心到树皮先增大而后减小, 这与基本密度的变化趋势基本一致(成俊卿, 1985)。从总体上说腐朽木材的平均干缩率基本上都略高于正常木。
2.6 腐朽木与正常木显微结构比较与扫描电镜分析首先制作普通切片在光学显微镜下观测比较腐朽木与正常木显微结构的差异。图版Ⅰ中图 1~8为圆盘照片及光学显微镜下拍摄的照片。然后制作喷金切片在扫描电镜下观测, 图版Ⅱ中图 1~12为扫描显微镜下拍摄的照片。从图版Ⅰ中图 4和图 1的比较看, 立木杨树树干腐朽有明显的夹皮现象。夹皮是指生活的树干受伤后, 以至这部分的树干因形成层死亡而停止生长, 但周围的组织仍然继续生长, 将树皮的一部分包进木材中所形成的一种缺陷, 有的夹皮形成后, 经过若干树木生长被包藏到木材的内部, 只有在横切木材或解锯时才发现, 称为内夹皮。内夹皮在树干的横断面上, 常形成包含树皮的弧形或环状裂隙。显露在树干外部, 从表面可以看见的夹皮称为外夹皮(成俊卿, 1985), 外夹皮在树干侧面常成沟状(图版Ⅱ-4)。夹皮使木材的正常生长发育受到破坏, 影响木材构造的均匀性。杨树易发生夹皮现象, 滩地上夹皮的杨树常常伴随着不同程度的腐朽, 这是由于夹皮产生裂隙使浸泡在水中的杨树更易遭受菌类的侵害所致, 夹皮是滩地杨树发生腐朽的直接原因, 但是生长在滩地的杨树是否更容易产生夹皮现象目前还不清楚, 亦即杨树夹皮现象的产生是受基因控制还是受外界环境控制目前还不清楚。从图版Ⅰ中图 2和图 5以及图版Ⅱ中图 1和图 4的比较看, 腐朽木横切面导管比量比正常木明显要低, 这使得腐朽木输导水分的功能减弱, 易造成菌类侵害。进一步放大观察, 从图版Ⅰ中图 3和图 6可见正常木纤维细胞形状十分规则整齐, 而腐朽木则出现应拉木区域, 其特征是纤维细胞出现明显的变形, 并且应拉木纤维胞壁上出现明显的胶质层(G层)(K.Wilson et al., 1986; 成俊卿, 1985), 胶质层是一层与细胞长轴呈5°的纤维素微纤丝外鞘, 其厚度常等于或大于正常胞壁的S 2层, 常见于靠近胞腔一边, 常起皱与膨大, 并局部分离, 如凝胶状填充在胞腔里, 由于胶质层(G层)的存在, 使应拉木纤维的胞壁比正常木的厚。图版Ⅰ图 6中颜色较深靠近胞腔一侧的为胶质层(G层), 进一步在扫描电镜下观测如图版Ⅱ图 7和图 8, 胶质层(G层)更加明显, 由于胶质层(G层)缺乏木质素使得在扫描电镜下呈浅白色。
图版Ⅱ中2、3及5、6分别为正常木早材和晚材的扫描照片, 可见晚材的胞壁率要高于早材, 即胞壁厚度要比早材厚, 同时晚材S2层相对比例比早材高, 从照片中可见晚材胞壁颜色较深的部分比早材多, 即晚材S2层木素含量比早材高。
图版Ⅰ中图 7和图 8均为腐朽木横切面腐朽区域, 在低倍横切面上可见很多小黑点是菌类侵害的结果, 高倍横切面上可见很多木纤维细胞被菌类侵染而呈深黑色, 但是细胞形状仍基本保持正常。进一步在扫描电镜下观测(见图版Ⅱ-9~12), 图 9腐朽木横切面清晰显示在导管和木纤维中均有菌丝分布, 图 10和图 11显示腐朽木径切面上菌丝的分布, 其中导管中发现大量菌丝如图 10, 其次木射线组织中也含有较多的菌丝如图 11, 而木纤维中所含菌丝相对较少, 这说明菌类侵害杨树可能首先入侵导管, 然后通过木射线细胞向木纤维细胞蔓延, 这至少说明导管很适宜菌丝的生存。图 12显示腐朽木弦切面上木纤维细胞内菌丝的分布, 菌丝在木纤维细胞间的蔓延主要通过纹孔传递扩散。
3 小结地势高低对杨树立木腐朽株数所占比例起决定作用, 地势低, 淹水时间长, 易使杨树腐朽株数增加, 一般淹水最高水位不超过树高的2.50 m的滩地种植杨树较适宜。
无论腐朽木还是正常木采用y =a·e-b/x描述杨树的径向生长即胸径与年龄之间的关系效果均较好, 相关系数较高。腐朽对杨树胸径生长变化趋势影响较小。腐朽木与正常木各面的年轮宽度、晚材率、含水率、基本密度、干缩率变化趋势一致, 均是自髓心到树皮先增大而后减小, 约在4-6 a达最大值, 而后略降低。
从外表看, 滩地上夹皮的杨树常常伴随着不同程度的树干腐朽, 夹皮是滩地杨树发生腐朽的直接原因。同时树干腐朽的杨树一般没有根腐现象, 说明菌类侵害立木杨树主要通过夹皮的裂隙。
腐朽对年轮宽度影响较小。腐朽材内部生材含水率低于正常材, 其内部生材含水率与外部生材含水率之差远低于正常材, 这说明腐朽材内部输导水分功能减弱, 与其导管比量减少相一致。腐朽材外部饱和含水率远低于正常材, 说明其吸水性较正常材弱。腐朽材的密度和干缩率比正常材略高, 主要由于腐朽材中通常出现应拉木区域, 应拉木区域木纤维细胞胞壁出现较厚的胶质层(G层)使得其密度高于正常材, 同时腐朽材导管比量比正常材低也是导致其密度高的原因。
正常材晚材的胞壁率要高于早材, 同时晚材S2层相对比例比早材高。
通过电镜观测发现侵害杨树的菌丝在导管中分布数量较多, 通过木射线细胞向木纤维细胞蔓延, 菌丝通过细胞间的纹孔进行扩散蔓延。
成俊卿. 1985. 木材学. 北京: 中国林业出版社.
|
陈敏忠, 王传槐, 等. 1996. 白腐菌云芝腐朽木的超微结构研究. 南京林业大学学报, (1): 48-52. |
丁佐龙, 费本华, 刘盛全. 1997. 木材白腐机理研究进展. 木材工业, (5): 18-21. |
江泽慧.兴林灭螺论文集.北京: 中国林业出版社, 1995
|
金重为, 邰瓞生, 尤纪雪. 1989. 天然耐腐木材的抗腐机理及其在腐朽过程中化学成份的变化. 林业科技, (5): 447-452. |
刘寿坡等.毛白杨胸径粗生长过程的观测.黄泛平原林地资源利用研究, 北京: 中国科学技术出版社, 1992, 33~27
|
彭镇华, 江泽慧. 1995. 中国新林种-抑螺防病林研究. 中国林业出版社. |
王宝松, 徐忠虞, 潘明建, 等. 1993. 杨树新无性系耐水性差异的研究. 江苏林业科技, (4): 1-4. |
王金林, 李春生, 陆从进. 1999. 杨木旋切及单板质量与木材性质关系的研究. 木材工业, 9(5): 1-7. |
曾其蕴, 等. 1990. 河北毛白杨木材纤维长度变异的研究. 林业科学, 26(3): 232-238. |
Bruce J. 1984. Zobel, John Talbert. New York: Applied Forest Tree Improvement, Wiley.
|
Bruce J.Zobel, Johannes P.van Buijtenen.Wood variation: its cause and control, Springer -Verlag, Berlin; New York, 1989
|
K.Wilson and D.J.B .White.The Anatomy of Wood:its diversity and variability.Stobart&Son Ltd, London, 1986
|