 2001, Vol. 37
2001, Vol. 37文章信息
- 陈少瑜, 吴丽圆, 李江文, 项伟, 周云.
- Chen Shaoyu, Wu Liyuan, Li Jiangwen, Xiang Wei, Zhou Yun.
- 云南红豆杉天然种群遗传多样性研究
- STUDY ON GENETIC DIVERSITY OF NATURAL POPULATIONS OF TAXUS YUNNANENSIS
- 林业科学, 2001, 37(5): 41-48.
- Scientia Silvae Sinicae, 2001, 37(5): 41-48.
-
文章历史
- 收稿日期:2000-01-10
-
作者相关文章


红豆杉属Taxus约有11种, 我国有4种1变种, 主要分布于东北、西南和华南(郑万钧等, 1978)。云南红豆杉(Taxus yunnanensis)便是其中在云南有分布且紫杉醇含量较高的一种1)。云南红豆杉在云南主要分布于哀牢山以西的湿润、半湿润季风气候区, 多散生于海拔2400 ~ 3200 m的高山台地、沟谷溪流两岸, 成为西南季风常绿阔叶林、针阔叶林、暗针叶林的组成部分(张茂钦等, 1996)。此树种能生产优良的木材, 也可作为庭院的美化树种, 但其更大的用途还在于80年代发现的药用价值, 它可作为既安全且有效的抗癌药———紫杉醇的提取原料(梁敬钰, 1993)。也正因为这一巨大的药用价值, 使得本来自然分布就很稀少的资源受到进一步的威胁。
1) 张茂钦等.珍稀濒危树种生物学特性及内含物检测研究报告.云南省林科所, 1992。
70年代以来, 国外广泛利用同工酶电泳技术研究树种的群体遗传结构, 取得了较满意的结果。如西黄松(Pinus ponderesa)(Linhart et al., 1981)、台地松(P. pungers)(Raymond et al., 1982)、辐射松(P. radiata)(Plessas et al., 1986)等。我国对马尾松(P. massoniana)、红松(P. koraiensis)、杉木(Cunninghamia lanceolata Hook)群体同工酶也作过报道(葛颂等, 1988; 扬一平等, 1989; 黄敏仁等, 1986)。目前, 对于云南红豆杉还缺乏此方面的研究。本项研究以滇西北三江流域的3个自然种群为分析对象, 通过等位酶电泳及遗传分析, 阐明其遗传多样性水平, 为合理保护和利用云南红豆杉基因资源提供基础资料和科学依据。
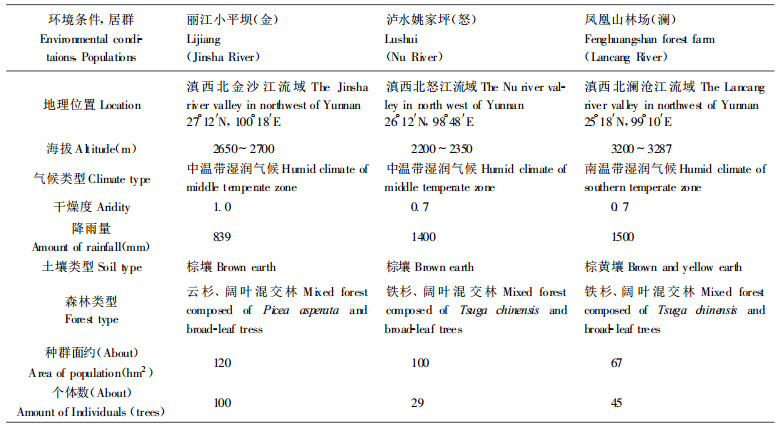
1 材料和方法 1.1 种子采集及电泳实验种子采于滇西北三江流域, 即怒江、澜沧江和金沙江流域的3个天然种群。由于云南红豆杉天然分布很稀少, 天然种群雌株数仅为雄株数的1 3强些, 个体间果实成熟早晚不一, 又加之人为破坏严重, 使得采种十分困难。每群体采种单株仅10株左右。采种后分单株脱粒、净种, 置于冰箱中保存备用。各居群的环境条件见表 1。
|
|
电泳采用聚丙烯酰胺凝胶垂直板状电泳, 不连续缓冲系统, 低温电泳并染色。以单粒胚乳为单位提取酶蛋白, 样品提取缓冲液为0.1mol Tris-HCl, (pH 7.5)+5 %蔗糖+0.1 %β巯基乙醇+10 % PVP(提取MDH, GOT, EST, SKD酶蛋白), 电极液+20 %蔗糖(pH8.3)(提取PER酶蛋白)。冰浴研磨, 10000 r min离心15 min, 取上清电泳。分离胶浓度为7.5 %, 浓缩胶浓度为3 %, 电极缓冲液为Tris-甘氨酸(pH 8.3)。电泳在4 ℃冰箱中进行, 浓缩胶部分电流1 mA样, 进入分离胶增至2 mA样。染色参照王仲仁(1996)、Cheliak et al. (1984)及扬一平等(1989)的方法进行。
1.2 等位酶分析分析的酶类为谷氨酸草酰乙酸转氨酶(Got, E.C. 2.6.1.1)、苹果酸脱氢酶(Mdh, E. C. 1.1.1.37)、酯酶(Est, E.C. 3.1.1.1)、莽草酸脱氢酶(Skd, E.C.1.11.11.25)和过氧化物酶(Per, E.C.1.11.1.7)。由于云南红豆杉种子的大配子体是单倍体, 分析单株树一定数量的雌配子体, 具多态位点的, 其同工酶表型分离应为1:1;具单态位点的, 其同工酶表型是一致的, 没有分离。这样便可直接从酶谱读出单株的基因型, 计算出各群体各基因位点的等位基因频率。(将杂合基因型错定为同质基因型的概率为0.5k-1, 其中k为分析的雌配子体数, 这样每个单株分析6 ~ 10个, 平均8个雌配子体同工酶的分离表型, 推测母树的基因型, 把杂合基因型错定为同质基因型的概率为0.007)(Boyle et al., 1987)。
1.3 遗传分析以各基因位点的等位基因频率为基本数据, 用以下遗传参数进行遗传多样性及遗传分化方面的研究(王仲仁, 1996; 庞广昌等, 1995)。
(1) 多态位点百分率P(Percentage of polymorphic loci)
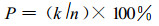
|
式中, K为多态酶位点的数目; n为所测定酶位点的总数。多态位点的标准(常用):最常见的等位基因出现频率小于或等于0.99的位点(Nei, 1975)。
(2) 每一位点平均观察的等位基因数A(Average number of alleles per locus)

|
式中, Ai为第i个位点上的等位基因数; n为所测定的位点。
(3) 平均每个位点等位基因的有效数Ae(Mean effective number of alleles per locus)
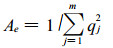
|
式中, qj为第j个等位基因的频率; m为测定到的等位基因的总数。
(4) 平均每个位点的观察杂合度Ho(Mean observed heterozygosity per locus)

|
式中, Hoi为第i个位点上的实际杂合度; qi第j个等位基因纯合基因型的频率; m为测定到的纯合基因型的总数; n为测定的位点总数; qij为第i个位点上第j个等位基因纯合基因型的频率; mI为第i个位点上的测定到的纯合基因型的种类数。
(5) 平均每个位点的预期杂合度He(Mean expected heterozygosity per locus)
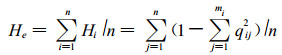
|
式中, Hi为第i个位点上的预期杂合度; n为测定的位点总数; qij为第i个位点上第j个等位基因的纯合基因型频率; mi为第i个位点上测到的等位基因总数。
(6) 各居群内基因多样度Hs(Gene diversity within population)

|
式中, m为i位点上的等位基因数; qi为i位点上第j个等位基因在亚居群中的频率。
(7) 总居群的基因多样度HT(Total gene diversity)
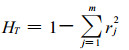
|
式中, m为i位点上的等位基因数; rj为i位点上第j个等位基因在总居群中的平均频率。
(8) 各居群间基因多样度DST(Gene diversity among populations)

|
(9) 基因分化系数GST(Proportion of interpopulation gene differentiation)

|
(10) 遗传距离D(Genetic distance)

|
式中, K为指定位点; Kk为X和Y两个居群在指定位点k上的遗传一致度; i为指定位点的等位基因; Xi为在居群X中位点k第i个等位基因的频率; Yi为在居群Y中位点k第i个等位基因的频率。
2 结果 2.1 群体遗传分析5个酶系统共出现14个位点, 遗传分析采用了其中各种群都出现且清晰的10个位点。其中Got-3, Per-2为单态位点, 其它Est-1, Est-3, Got-1, Got-2, Mdh-1, Mdh-4, Skd-1和Skd-2 8个位点为多态位点。(多态位点的标准是最常见等位基因的频率≤0.99)Est-1位点有1条或2条带的不同的4个等位基因, Est-3显示2个不同迁移率的双带, GOT的两个位点皆表现为2个单带的等位基因, Mdh- 1位点有3个不同迁移率的双带, 显示3个等位基因, Mdh-4位点出现了“转录后饰变”现象, 即出现3 ~ 4条相互间不杂合的平行带, 表现为2个等位基因, Skd-1位点有1 ~ 2条带的4个等位基因, Skd-2则显示了2个单带的等位基因。包括同质位点的基因, 共有25个等位基因。5种酶的带型见图 1。5种酶的电泳酶谱式样见图 2~6。

|
图 1 云南红豆杉等位酶图谱 Fig. 1 Schematic zymographic patterns of five enzymes in female gametophte of Taxus yunnanensis |

|
图 2 SKD 2个位点的电泳酶谱式样 Fig. 2 Banding pattern of two loci of SKD |

|
图 3 EST 3个位点的电泳酶谱式样 Fig. 3 Banding pattern of three loci of EST |

|
图 4 MDH 4个位点的电泳酶谱式样 Fig. 4 Banding pattern of four loci of MDH |

|
图 5 GOT 3个位点的电泳酶谱式样 Fig. 5 Banding pattern of three loci of GOT |

|
图 6 PER 2个位点的电泳酶谱式样 Fig. 6 Banding pattern of two loci of PER |
群体的遗传多样性由遗传参数P, A, Ae, He, Ho描述。各遗传参数的计算公式见前述, 其计算的基本数据等位基因频率(Allele frequency)的计算公式为
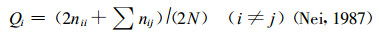
|
式中, nii为具有纯合aiai基因型的个体数; nij为具有杂合aiaj基因型的个体数; N为个体总数。
5个酶系统10个基因位点(包括单态位点)等位基因频率和杂合性见表 2。
|
|
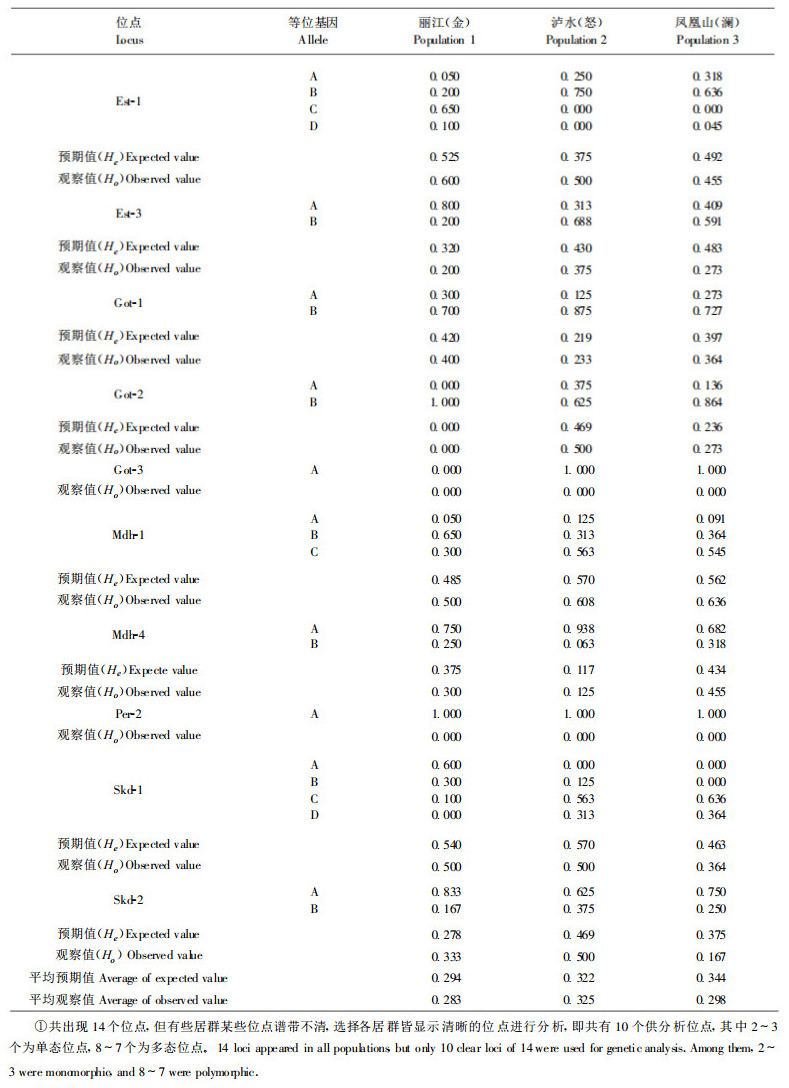
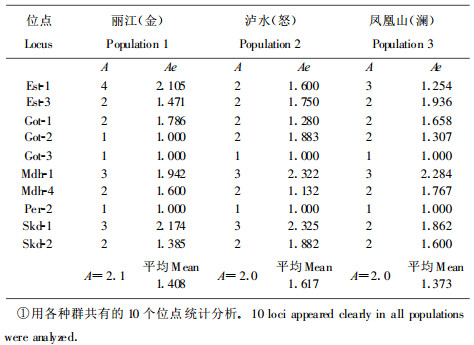
从表 2中可以看出, 群体间无论等位基因频率或杂合性均存在着差异。等位基因频率大的, 在各群体中均为主导; 等位基因小的, 在各群体中也低。杂合性的观察值变动范围为0.283 ~ 0.325, 其平均观察值(Ho)为0.299。杂合性的预期值的变动范围为0.291 ~ 0.344, 其平均值(He)为0.313。平均杂合性的观察值与预期值很接近(仅相差0.014)。各居群每个位点的多态位点比例, 平均每个位点的等位基因数及有效等位基因数见表 3。综上所述, 度量云南红豆杉群体遗传多样性丰富程度的各项数值列于表 4。
|
|
|
|

群体间的遗传分化状况由遗传参数HT, HS, DST, GST, D描述。具体的计算公式见前述。云南红豆杉分析群体间的遗传分化状况见表 5。
|
|
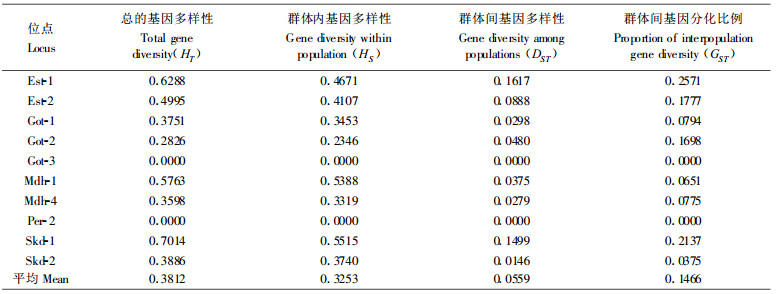
由表 5可以看出整个群体总的基因多样性(HT)平均值为0.3812, 群体内基因多样性(HS)平均值为0.3253, 群体间基因多样性(DST)平均值为0.0559, 群体间基因分化比例(GST)平均值为0.1466。群体间基因分化的相对值约15 %。换言之, 总的基因多样性85 %以上产生在群体内。
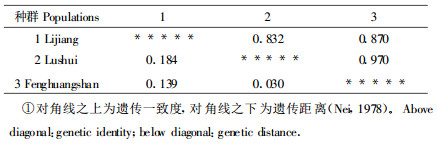
群体间的遗传距离矩阵见表 6。所有群体间的平均遗传距离为0.118。
|
|
据Hamrick和Godt对165属449种植物的统计结果, 植物种群内平均多态位点比率为34 %, 平均期望杂合度为0.11;他们对55个裸子植物物种的多态位点百分率, 有效等位基因数及平均杂合率等遗传参数作过总结, 结果是P =70. 9 %, Ae =2. 35, He =0. 173(Hamrick et al., 1989), 葛颂也总结过25个针叶树种的研究结果(葛颂, 1988), P =61. 5 %, Ae =2. 26, He =0. 206。分析的3个云南红豆杉天然群体的P, Ae, Ne的平均值分别为P =0. 75, Ae =1. 499, He =0. 312, 与以上统计结果相比较, 云南红豆杉天然群体的遗传多样性水平是比较高的。从表 5可以看出, 云南红豆杉群体内的平均基因多样性(HS)是0.3253, 比群体间的平均多样性(DsT =0.0559)大得多。群体间的基因分化系数(GST)是0.1466, 即只有大约总基因多样性的15 %是属于群体间基因差异, 而总基因多样性的约85 %产生于群体内。以上结果表明群体内的遗传多样性比群体间的遗传多样性大得多。
云南红豆杉群体间存在一定的遗传分化。据Hamrick (Hamrick et al., 1989)对91个植物物种的总结, GST =0.206;其中15个松科树种的GST =0.061。本次研究的GST =0.1466, 其值低于91个物种的平均值, 却高于15个松科树种的平均值。用遗传距离(D)也可表示群体间相对遗传分化的大小, 本研究的遗传距离平均值为0.118, 也说明了群体间存在一定的分化。另外, I与D值还表明各群体间的亲缘关系, 由表 6可看出, 泸水和凤凰山种群间的亲缘关系最近(I =0. 970, D =0. 03), 泸水和丽江种群间的亲缘关系最远(I =0. 832, D =0. 184)。一般说, 就遗传一致度的值来看, 植物属内种间的为0.67, 而种内居群间的为0.90(Hamrick et al., 1990)。云南红豆杉3个种群的I值接近于种内居群间的0.90这个标准, 说明各天然种群在遗传结构上的差异很小, 亲缘关系较近。如果种群间的空间距离加大, 相互的基因流便会减小, 内繁育机会增大, 使得种群间的遗传分化加大。Heywood(1991)曾指出, 种群间的遗传距离往往和它们的空间距离呈正相关。本研究结果也表明各种群间遗传分化的大小与空间距离成正相关。
云南红豆杉群体内的遗传多样性比较高, 主要原因是寿命长, 世代重叠, 其林分包括几个龄级, 不同龄级在建立时, 都会遇到当时的选择压力, 适宜的基因就保留下来, 因此积累了较多的基因, 形成了较广的遗传基础。另外, 云南红豆杉在云南的分布表现出较广的垂直分布(海拔1647 ~ 3300 m), 跨越了几个气候带(张茂钦, 1998), 这样一个较大的气候变化范围, 对不同种群的等位基因造成不同的压力, 因而在维持种群遗传多样性上也起到了一定作用。
云南红豆杉为雌雄异株, 风媒受粉, 其自然分布多为散生或群状散生, 极少成纯林, 并且成不连续分布, 因此种群间有一定分化。但由于此树种具有很强的传粉能力及结实能力(张茂钦, 1998), 带甜味的肉质假种皮助于种子的传播, 加之种群间没有阻止基因流的严重障碍, 导致种群间的遗传分化较低。
以上研究结果将对云南红豆杉基因资源的保护及开发利用策略的制定提供基础资料和科学依据。
葛颂, 等. 1988. 用同工酶研究马尾松群体的遗传结构. 林业科学, 24(4): 399-408. |
葛颂. 1988. 同工酶与林木群体遗传变异研究. 南京林业大学学报, (1): 68-75. |
黄敏仁, 等. 1986. 杉木种源酯酶同工酶地理分布研究. 南京林业大学学报, (3): 31-35. |
梁敬钰. 1993. 国外紫杉醇研究进展. 生物工程进展, 13(4): 19-20. |
庞广昌, 姜冬梅. 1995. 群体遗传多样性和数据分析. 林业科学, 31(6): 543-550. DOI:10.3321/j.issn:1001-7488.1995.06.002 |
王仲仁. 1996. 植物等位酶分析. 北京: 科学出版社.
|
杨一平, 等. 1989. 红松群体内和群体间同工酶变异的研究. 林业科学, 25(3): 201-208. DOI:10.3321/j.issn:1001-7488.1989.03.002 |
张茂钦, 李达孝, 等. 1996. 云南红豆杉人工栽培及其生态生物学特性研究. 林业科技通讯, (3): 8-11. |
张茂钦. 1998. 云南珍稀濒危树种生态生物学研究. 昆明: 云南大学出版社.
|
郑万钧, 傅国立编. 1978. 中国植物志. (第七卷). 北京: 科学出版社.
|
Boyle T J B, Morgenstern E K. 1987. Some aspects of population structure of black spruce in central New Brunswick. Silvae Genet, 7(36): 53-60. |
Cheliak W M, Pitel J A. 1984. Techniques for Starch Electrophoresis of Enzymes from Forest Tree Species. Petawawa Natal.For.Inst.(Inf.Rep.PIX-42): 21-384. |
Hamrick J L, Golt M J.Allozyme diversity in plant species.In: Brown A H D, M T Clegg, A L Kahler et al..Plant population genetic, breeding, and genetic resources.Sunderland: Sinauer Associates, 1989, 43~ 63
|
Hamrick J L, Godt M J W. 1990. Allozyme diversity in plant species. Sunderland, Mass:Sinauer: 43-63. |
Heywood J S. 1991. Spatial analysis of genetic variation in plant population. Annals Review of Ecology System, 22: 335-355. DOI:10.1146/annurev.es.22.110191.002003 |
Linhart Y B et al.In: proc.Sym.Isozymes of North American Forest Trees and Forest Insect, 1981, 53~ 59
|
Nei M. 1987. Molecular evolutionary geneeics. New York: Columbia Univ.Press.
|
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590. |
Plessas M E and Strauss S H.Can.J.For.Res., 1986, 16: 1155~ 1164
|
Raymond P Guries and F Thomas Leding.Evolution, 1982, 36(2): 387~ 402
|