 2001, Vol. 37
2001, Vol. 37文章信息
- 张彦广, 王志刚, 黄大庄.
- Zhang Yanguang, Wang Zhigang, Huang Dazhuang.
- 桑天牛羧酸酯酶和谷胱甘肽S-转移酶与杨树次生代谢物质相关性研究
- INTERRELATION BETWEEN CARBOXYLESTERASE AND GLUTATHIONE-S-TRANSFERASE IN APRIONA GERMARI LARVAE AND SECONDARY METABOLITES OF POPLAR TREES
- 林业科学, 2001, 37(3): 106-111.
- Scientia Silvae Sinicae, 2001, 37(3): 106-111.
-
文章历史
- 收稿日期:2000-01-20
-
作者相关文章


解毒酶是昆虫体内一些能够代谢有毒外源化合物的酶类, 如羧酸酯酶、谷胱甘肽转移酶等, 能使昆虫自身减少或免除毒害作用。植物次生代谢物质是植物体内复杂的分支代谢途径的最后产物, 如酚类、萜类、生物碱类等, 它们不直接参与维持产生者的生长发育和生殖有关的原始生化活动。对于昆虫, 它们是非营养性物质; 对于植物, 它们具有对昆虫进行化学防御的生态功能。许多次生代谢物质如果随昆虫取食而在体内积累, 或延缓昆虫生长发育, 或降低昆虫繁殖率, 甚至会导致昆虫死亡。但昆虫能够在自己的寄主植物上顺利生长发育, 解毒酶的作用是不可忽视的一个重要因素(钦俊德, 1987)。
植物次生代谢物质对昆虫具有防御功能, 昆虫也能够依靠解毒酶的作用和其它一些解毒功能克服寄主植物次生代谢物质障碍。昆虫在取食不同寄主植物时, 还能够诱导体内解毒酶活力发生改变。对烟芽夜蛾(Heliothis virescens) (Riskallah et al., 1986), 虎凤蝶(Papilio glaucus) (Lindroth, 1989), 棉铃虫Helicoverpa armigera (谭维嘉等, 1990)、棉蚜Aphis gossypii (高希武, 1992)、小菜蛾Plutella xylostella (李云寿等, 1996)等昆虫的研究, 发现这些昆虫取食不同种类的植物时, 其体内的解毒酶活力会发生明显改变, 并认为这种改变是由于植物体内次生代谢物质的诱导作用导致的。
桑天牛Apriona germari寄主范围广泛, 对多种林木危害十分严重, 尤其以广泛栽培的毛白杨受害最为严重。选育抗天牛杨树品种是杨树天牛综合治理研究的重要内容, 了解树木对天牛的化学防御机制是一个重要方面。关于杨树化学成分(营养物质和次生代谢物质)与抗天牛关系方面的研究已有一些报道(杨雪彦等, 1992;王克胜等, 1995;王蕤等, 1995;孙丽艳等, 1995;黄大庄等, 1996)。这些研究多以分析杨树化学成分和调查杨树被害率的方法来研究两者的关系, 并依此确定哪些化学成分与抗天牛有关。多数结论认为, 杨树某些酚类物质的含量差异与杨树被害率相关联。
本研究以被认为能够影响昆虫生理代谢的植物次生代谢物质, 及其对昆虫生理代谢(解毒酶活力)的影响程度为主要内容, 选用不同的杨树作为桑天牛幼虫的取食对象, 研究取食不同杨树的桑天牛幼虫体内解毒酶活力是否发生改变, 分析不同杨树的次生代谢物质种类和含量存在的差异, 探讨取食不同杨树的桑天牛幼虫解毒酶活力与杨树次生代谢物质种类的相关性。目的是探明影响虫体生理代谢的次生代谢物质种类, 为阐明杨树抗桑天牛的机理, 确定杨树抗桑天牛的化学指标, 选育抗桑天牛杨树品种提供依据。
1 材料与方法 1.1 杨树的选用选用以下杨树作为桑天牛幼虫的取食对象:毛白杨(Populus tomentosa)、新疆杨(P.alba var.pyramidalis), I-214杨(P.×euraericana cv.) “I-214”、加拿大杨(P.canadensis)、山海关杨(P.canadensis cv.“Shanhaiguanensis”), 北京杨(P.×beijingensis), J-1杨*, J-2杨* (*为正在选育的品种)。
1.2 桑天牛的饲养田间捕捉成虫, 放入养虫笼内以桑树枝条饲养。成虫交配并产卵于枝条内, 每日更换枝条, 从中剖取当日虫卵放在培养皿中并保持一定湿度(底部垫湿润脱脂棉, 上衬一层滤纸, 虫卵置于滤纸上面。在虫卵孵化当日, 将小幼虫接种于以上8种杨树2年生枝条的人工刻槽内。幼虫取食10d, 从枝条内剖取幼虫, 用于测定解毒酶活力。
1.3 桑天牛幼虫解毒酶活力的测定 1.3.1 酶源制备桑天牛幼虫5head/Group, 放入玻璃匀浆器, 加水2.5mL, 冰浴下匀浆。4℃, 10000r/min, 离心30min, 取上清液作为酶源(2头/mL)。取食8种杨树的幼虫各制备3份样品作为3次重复。
1.3.2 酶源蛋白质含量测定反应体系为0.01head/mL酶源0.4mL, 水2.6mL, 0.02% CBB-G250 3.0mL总体积6mL。混匀后, 用分光光度计测595nm处光密度值, 重复3次, 根据蛋白质标准曲线方程求酶源蛋白质含量。
1.3.3 羧酸酯酶酶活力测定反应体系为0.04mol/L pH7.0磷酸缓冲液0.7mL, 0.0003mol/L醋酸-α-萘酯底物液(含0.0001mol/L水杨酸毒扁豆碱) 5mL, 0.01head/mL酶源0.3mL, 总体积6mL。在37℃水浴中振荡30min, 加入坚牢蓝B显色剂1mL, 25℃恒温放置30min后分光光度计测600nm处光密度值。根据α-萘酚标准曲线方程求出酶促生成的α-萘酚量, 酶的比活力表达为α-萘酚mmol/L/mg pr./30min。使用SAS统计分析软件对酶的比活力进行方差分析。
1.3.4 谷胱甘肽S-转移酶活力测定反应体系为0.066mol/L pH7.0磷酸缓冲液(含0.002mol/L乙二胺四乙酸) 2.5mL, 0.05mol/L还原型谷胱甘肽底物液0.3mL, 0.03mol/L 2, 4-二硝基氯苯丙酮底物液0.1mL, 1head/mL酶源0.1mL, 总体积3mL。25℃下, 用分光光度计测反应5min的340nm处光密度值。酶的比活力表达为OD值/mg pr./5min。使用SAS统计分析软件对酶的比活力进行方差分析。
1.4 杨树次生代谢物质分析 1.4.1 杨树次生代谢物质种类酚类有苯酚、邻苯二酚等; 酚酸类有苯甲酸、水杨酸、对羟基苯甲酸、儿茶酸、没食子酸、丁香酸、肉桂酸、香豆酸、咖啡酸、阿魏酸、绿原酸等; 甙类有水杨甙。
1.4.2 样品制备取选用的8种杨树2年生枝条, 剥去韧皮部, 将木质部剪碎并烘干, 粉碎后过40目筛, 120℃烘至恒重。
酚类 干粉2.0000g放于三角瓶中, 加50mL石油醚, 超声波振荡2h, 放置过夜, 滤去石油醚。将残渣晾干, 转入烧瓶中, 加40%甲醇50mL, 加热回流2h, 过滤, 滤液用甲醇定容至50mL, 摇匀, 经0.5μm滤膜过滤, 清液待测定。
酚酸类 干粉3.0000g放于三角瓶中, 加50mL石油醚, 超声波振荡30min, 放置过夜, 滤去石油醚。将残渣晾干, 转入烧瓶中, 加50%甲醇50mL, 加热回流1h, 过滤, 滤液浓缩定容至25mL。取Waters公司Sep-pak C18小柱1支, 用5mL甲醇冲洗活化, 再用5mL水冲洗。吸取5mL样品提取液通过小柱, 弃去最初洗出液1mL, 再收集2mL洗出液待测定。
水杨甙 干粉5.0000g放于三角瓶中, 加30mL乙醚过夜脱脂。滤去乙醚, 残渣中加50%甲醇50mL, 加热回流1h, 过滤, 滤液中加10%醋酸铅10mL, 摇动2min, 滤去沉淀物, 滤液浓缩至干, 用甲醇重新溶解并定容至50mL, 摇匀, 经0.5μm滤膜过滤, 清液待测定。
1.4.3 次生代谢物质含量测定采用高效液相色谱分析法, 仪器为Waters 244型HPLC, Kromasil C18柱(0.4cm×15cm)。定量方法采用外标法(峰高)定量, 洗脱速度0.7mL/min。
色谱条件:
① 苯酚、苯甲酸、水杨酸、肉桂酸。流动相:65%甲醇, 35%水, 0.1%磷酸调pH=4。检测器:UV 232nm×0.1 AUFS。
② 邻苯二酚、对羟基苯甲酸、儿茶酸、没食子酸、丁香酸、香豆酸、咖啡酸、阿魏酸、绿原酸。流动相:30%甲醇, 70%水, 0.1%磷酸调pH=4。检测器:UV 254nm×0.1 AUFS。
③ 水杨甙。流动相:40%甲醇, 60%水, 0.1%磷酸调pH=4。检测器:UV 254nm×0.1 AUFS。
使用SAS统计分析软件对次生代谢物质含量测定结果进行方差分析和聚类分析。
1.4.4 标样和试剂苯酚、邻苯二酚、苯甲酸、水杨酸、对羟基苯甲酸、儿茶酸、没食子酸、丁香酸、肉桂酸、香豆酸、咖啡酸、阿魏酸、绿原酸:色谱纯(Sigma公司)。
水杨甙:分析纯(北京化工厂)。水:重蒸, 并经0.45μm滤膜过滤。甲醇:优级纯, 并经0.5μm滤膜过滤。磷酸:优级纯。石油醚、乙醚、乙醇、醋酸铅:分析纯
1.5 酶活力与次生代谢物质逐步回归分析使用SAS统计分析软件对酶的比活力与次生代谢物质进行逐步回归分析。
2 结果与分析 2.1 羧酸酯酶活力测定结果羧酸酯酶比活力的大小顺序为山海关杨 > 加拿大杨 > 新疆杨 > 毛白杨 > 北京杨 > I-214杨 > J-2杨 > J-1杨。方差分析表明, 取食不同杨树的桑天牛幼虫羧酸酯酶活力存在显著差异。由多重比较看到, 山海关杨与J-2杨、J-1杨; 加拿大杨与J-1杨之间存在显著差异(见表 1)。
|
|

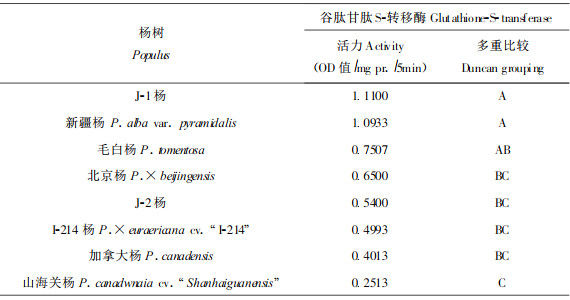
谷胱甘肽S-转移酶比活力的大小顺序为J-1杨 > 新疆杨 > 毛白杨 > 北京杨 > J-2杨 > I-214杨 > 加拿大杨 > 山海关杨。方差分析表明, 取食不同杨树的桑天牛幼虫谷胱甘肽S-转移酶活力存在显著差异。由多重比较看到, J-1杨、新疆杨、毛白杨与山海关杨; J-1杨、新疆杨与北京杨、J-2杨、I-214杨、加拿大杨之间存在显著差异(见表 2)。
|
|
8种杨树枝条木质部次生代谢物质含量测定结果见表 3。从分析的14种次生代谢物质来看, 8种杨树均缺少苯甲酸、儿茶酸、没食子酸、咖啡酸; 此外, 新疆杨缺少邻苯二酚、香豆酸、阿魏酸; 加拿大杨缺少苯酚、肉桂酸、香豆酸、阿魏酸; I-214杨和山海关杨缺少肉桂酸、香豆酸; 北京杨缺少苯酚、香豆酸; J-1杨缺少肉桂酸、香豆酸、阿魏酸; J-2杨缺少苯酚、肉桂酸、香豆酸、阿魏酸。
|
|

8种杨树枝条木质部次生代谢物质含量多重比较结果见表 3。方差分析表明, 8种杨树所含10种次生代谢物质的含量存在显著差异。
8种杨树枝条木质部次生代谢物质含量聚类分析结果见表 4。类平均法聚类分析表明, 当取平均距离阈值T=35时, 可将8种杨树分为{加拿大杨、I-214杨、北京杨、J-2杨、J-1杨、山海关杨}与{毛白杨、新疆杨}2类。
|
|

羧酸酯酶与次生代谢物质逐步回归分析表明, 有苯酚(x1)、对羟基苯甲酸(x5), 丁香酸(x8), 绿原酸(x13) 4种次生代谢物质进入逐步回归方程。其中苯酚(x1)是加拿大杨、北京杨所缺少的种类。羧酸酯酶与次生代谢物质逐步回归方程:y=87.1935-2.2423x1-2.3059x5+24.2697x8+1.0837x13
谷胱甘肽S-转移酶与次生代谢物质逐步回归分析表明, 有苯酚(x1), 阿魏酸(x12) 2种次生代谢物质进入逐步回归方程。其中苯酚(x1)是加拿大杨、北京杨所缺少的种类; 阿魏酸(x12)是新疆杨、加拿大杨、J-1杨、J-2杨所缺少的种类。谷胱甘肽S-转移酶与次生代谢物质逐步回归方程:
y=0.5329+0.0258x1-0.0724x12
3 讨论 3.1 关于不同杨树对解毒酶活力的影响昆虫取食不同植物诱导解毒酶活力发生改变的现象, 除了鳞翅目的棉铃虫、小菜蛾等和同翅目的棉蚜等农业重要害虫(谭维嘉等, 1990;高希武, 1992;李云寿等, 1996)以外, 在鞘翅目的桑天牛这一林业重要害虫也得到了证实。方差分析表明, 取食不同杨树的桑天牛幼虫羧酸酯酶和谷胱甘肽S-转移酶活力存在显著差异。从多重比较中看到, 两种解毒酶活力受到取食对象的影响有所不同。因此可以认为, 取食不同的杨树对两种解毒酶活力影响的内在因素是不同的。仅就本研究探讨的次生代谢物质方面来看, 诱导两种解毒酶活力发生改变的次生代谢物质种类可能是不同的。
3.2 关于不同杨树次生代谢物质种类和含量的差异高效液相色谱分析表明, 不同杨树的次生代谢物质种类有所不同。方差分析表明, 不同杨树的不同次生代谢物质在含量上也存在差异。从多重比较的结果来看, 在不同杨树间各次生代谢物质含量的差异情况比较复杂, 这暗示杨树次生代谢物质对昆虫生理代谢的影响也会是复杂的。这种复杂的差异可能是导致解毒酶活力存在差异的原因之一。
对8种杨树枝条木质部次生代谢物质含量进行聚类分析, 可将8种杨树分为{加拿大杨、I-214杨、北京杨、J-2杨、J-1杨、山海关杨}与{毛白杨、新疆杨}2类。在我国北方恰以白杨派的毛白杨、新疆杨受桑天牛危害严重, 而较少危害黑杨派杨树(阎浚杰等, 1999)。两类群杨树在某些次生代谢物质上存在的差异, 是否可以做为对桑天牛的抗性化学指标, 有待进一步研究。
3.3 关于解毒酶活力与次生代谢物质的相关性研究发现, 对羧酸酯酶有显著影响的次生代谢物质种类有4种, 对谷胱甘肽S-转移酶有显著影响的次生代谢物质种类有2种, 其中苯酚是共同的, 其它则是不同的。从逐步回归方程可以看到, 如果杨树丁香酸、绿原酸含量增高, 苯酚、对羟基苯甲酸含量降低, 则羧酸酯酶活力会较高; 如果杨树苯酚含量增高, 阿魏酸含量降低, 则谷胱甘肽S-转移酶活力会较高。其中苯酚含量对两种解毒酶活力的影响是相反的。在选育抗桑天牛杨树品种时, 这种相关性及其影响方式可以做为分析抗虫机制的参考依据。
高希武. 1992. 寄主植物对棉蚜羧酸酯酶活性的影响. 昆虫学报, 35(3): 267-272. DOI:10.3321/j.issn:0454-6296.1992.03.021 |
黄大庄, 等. 1996. 树木单宁含量和抗桑天牛的关系. 河北省科学院学报, 13(3): 293-295. |
李云寿, 等. 1996. 不同寄主植物对小菜蛾羧酸酯酶活性的影响. 山东农业大学学报, 27(2): 147-151. |
钦俊德. 1987. 昆虫与植物的关系. 北京: 科学出版社.
|
孙丽艳, 等. 1995. 对云斑天牛有不同抗性的杨树品种中化学物质的分析. 林业科学, 31(4): 338-345. |
谭维嘉, 等. 1990. 取食不同寄主植物的棉铃虫对溴氰菊酯敏感性的变化. 昆虫学报, 33(2): 155-160. DOI:10.3321/j.issn:0454-6296.1990.02.019 |
王克胜, 等. 1995. 杨树抗云斑天牛纤维材无性系选育. 林业科学研究, 8(4): 429-436. DOI:10.3321/j.issn:1001-1498.1995.04.014 |
王蕤, 等. 1995. 毛白杨树皮内含物对光肩星天牛抗生的探讨. 林业科学, 31(2): 185-188. DOI:10.3321/j.issn:1001-7488.1995.02.001 |
杨雪彦, 等. 1992. 不同杨树营养物质对黄斑星天牛抗性的研究. 西北林学院学报, 7(3): 26-32. |
阎浚杰, 等. 1999. 光肩星天牛生态控制模式的研究. 河北农业大学学报, 22(4): 83-87. |
Lindroth R L. 1989. Host plant alteration of detoxication activity in Papilio glaucus L. Entomol.Exp.Appl, 50: 29-35. DOI:10.1111/j.1570-7458.1989.tb02310.x |
Riskallah M R, et al. 1986. Host plant induction of microsomal monooxygenase activity in relation to diazinon metabolism and toxicity in larvae of the tobacco budworm Heliothis virescens(F.). Pestic.Biochem.Physiol, 25: 233-247. DOI:10.1016/0048-3575(86)90050-7 |