 2001, Vol. 37
2001, Vol. 37文章信息
- 张飞萍, 蔡秋锦, 钟景辉.
- Zhang Feiping, Cai Qiujin, Zhong Jinghui.
- 竹缺爪螨实验种群生命表的研究
- STUDY ON POPULATION LIFE TABLE OF APONYCHUS CORPUZAE RIMANDO
- 林业科学, 2001, 37(3): 101-105.
- Scientia Silvae Sinicae, 2001, 37(3): 101-105.
-
文章历史
- 收稿日期:2000-01-18
-
作者相关文章


2. 泉州市林业局森防站 泉州 362000
2. Quanzhou Bureau of Forestry Quanzhou 362000
竹缺爪螨(Aponychus corpuzae Rimando)可取食多种竹子叶片, 在毛竹(Phyllostachys heterocycla cv.pubescens)和黄金间碧竹(Bambusa vulgaris cv.vittata)上造成危害。蔡秋锦等(1999)就其生物学及其发生与环境的关系作了报道, 但有关种群生命表研究未见报道, 传统的生命表分析方法应用在螨类研究中存在较大的误差且未能表达更多的信息, 冯士雍(1982)、杨凤翔等(1991)认为应用生存分析理论能够较好地表达种群的生存状态。Hsin Chi(1985, 1988)应用多维矩阵研究昆虫种群生命表具有较大的优点, 但螨类的发育过程与昆虫有较大区别。本文应用这两种方法分析了25℃下竹缺爪螨在黄金间碧竹叶片上取食种群变动规律并就两种方法在分析螨类种群生命表中的应用作了讨论。
1 材料与方法 1.1 试验方法1999年5月从黄金间碧竹叶上采回大量雌成螨, 另取部分叶片用棉花拭擦干净后于双目解剖镜下检查叶正、反面均无它物方可, 将采回雌成螨置于叶片背面待其产卵, 次日8:00检查, 将所产卵当作当日产卵置于25℃恒温培养箱中饲养(蔡秋锦等, 1999), 每日8:00观察卵孵化情况, 孵化后将幼螨移至另一叶上(单叶单螨)编号于培养皿中饲养, 每日早8:00~10:00时检查并记录各螨存活、生长发育及繁殖的情况, 试验总数112头。
1.2 分析方法 1.2.1 生存分析理论(冯士雍, 1982;杨凤翔, 1991)以3d为一年龄阶段将竹缺爪螨发育年龄分为[ti-1, ti]i个区间, 则实际划分区间数i=42/3=14个; 区间中点tmi=(ti-ti-1)/2, 区间长度hi=ti-ti-1; 设n′i为进入该区间总螨数, li为该区间内因浸水, 换叶和失踪造成死亡数, 则区间内校正螨数ni=n′i-li/2;再设di为区间内的自然死亡数, 则种群各生存参数可由下列方法估算(生命表格式见表 1) : 死亡率





如图 1, 图中A矩阵为种群年龄—阶段结构矩阵, nij为特定年龄—阶段个体数, i为年龄(单位:d), 以天为一年龄阶段, 则i=1, 2, …, 42;j为发育阶段, j=1, 2, 3, 4, 5, 6;分别指卵、幼螨、前若螨、后若螨、雄成螨、雌成螨(下同); B为种群稳定状态下特定年龄—阶段分布矩阵, mij为第i龄第j发育期个体占总螨数百分率, 
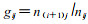






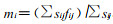
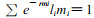

|
图 1 多维矩阵组建流程 Fig. 1 Construction course of multidimensional matrix |
根据生存分析理论以3d为年龄阶段组建竹缺爪螨25℃下实验种群生命表(表 1), 由此绘制死亡密度函数曲线和危险率函数曲线(图 1)。由表 1、图 2可知, 

|
|


|
图 2 死亡密度函数和危险率函数曲线 Fig. 2 Function curves of mortality density and of hazard ratio
—◆—死亡密度(tmi) mortality density (tmi); —■—危险率( Hazard ratio Hazard ratio 
|
根据年龄—阶段结构矩阵(A矩阵)组建存活率S矩阵(表 2), 由S矩阵绘制特定年龄—阶段存活率曲线, 如图 3, 曲线表明, 竹缺爪螨各发育期间存在明显的重叠现象, 5~9d内卵与幼螨, 前若螨重叠, 幼螨和前若螨则可与任何螨态重叠, 后若螨、雄成螨、雌成螨之间均能重叠出现且都能与幼螨、前若螨重叠, 其集中重叠时间为11~13d期间, 19d后只出现成螨。曲线还清楚地表明各螨态存在高峰期, 幼螨存在高峰期为6~9d, 前若螨为9~13d, 后若螨为13~16d, 雌雄成螨为16~27d。
|
|


|
图 3 竹缺爪螨年龄-阶段存活曲线 Fig. 3 Age-stage survival curves of A.corpuzae —◆—卵Egg, —■—幼虫Larv, —△—前若虫Protonymph, —+—后若虫Deutonymph, —*—前雄成虫Adult Male, —●—雌成虫Adult femal |
由表 2 S矩阵、F矩阵计算出在25℃, 以黄金间碧竹为食物人工饲养条件下, 具有稳定年龄组配的竹缺爪螨种群最大瞬间增长速率(内禀增长率) rm=0.0811, 周限增长率λ=1.0845/d, 每1世代种群增殖倍数(净增殖率) Ro=5.8368, 平均世代长度T=21.7532d。
2.4 竹缺爪螨种群稳定的年龄—阶段分布由A矩阵经计算建立稳定年龄—发育期分布矩阵(B矩阵)如表 3, 表中横行之和为某年龄在稳定年龄分布中所占百分比, 纵行之和为某发育期在稳定发育期分布中所占百分比。竹缺爪螨种群在实验室条件下达到稳定状态后卵和成螨最多, 分别占有27.55%和27.79%, 后若螨最少, 仅占6.37%。如果将竹缺爪螨年龄分为1~10d, 11~20d, 1~30d, 31d以后4组, 则各年龄段在种群稳定条件下各占有44.84%、31.44%、18.5%、5.22%。
|
|

对于体小质软, 人工饲养较困难的螨类, 在饲养过程中常因触水、换叶或失踪(如竹缺爪螨)死亡而导致试验中断或样本数大量减少, 应用生存分析理论分析其种群生存过程, 能够充分利用已得数据, 同时又能够较为准确地估算出其Ŝ(ti)、


Hsin Chi等(1985)多维矩阵模型用于分析种群生命表能够有效地减少个体发育间差异。既考虑了不同发育阶段重叠所带来的误差, 同时又考虑了雌雄个体不同的发育过程, 将它应用在重叠现象严重, 雌雄个体发育过程差异大的竹缺爪螨种群变动分析上, 颇具优点, 还能清楚地描述各发育阶段的起始时间、数量和生存情况、重叠情况以及稳定状态下特定年龄—发育阶段的稳定分布, 但由于螨类发育过程的特殊性, 矩阵变换公式应做相应改动。
竹缺爪螨在不同营养、不同环境条件下发育历期及其性比等均有差异(蔡秋锦等, 1999)。文中仅讨论了25℃下利用黄金间碧竹叶片饲养种群的变动情况, 要获得更加全面的认识, 需要在不同的生态条件下进行系列研究。
蔡秋锦, 张飞萍, 钟景辉, 等. 1999. 竹缺爪螨生物学特性及其发生与环境的关系. 林业科学, 35(6): 76-80. DOI:10.3321/j.issn:1001-7488.1999.06.012 |
丁岩钦. 1994. 昆虫数学生态学. 北京: 科技出版社, 175-178.
|
冯士雍. 1982. 生存分析(Ⅰ). 数学的实践与认识, (3): 72-80. |
杨凤翔, 王顺庆, 徐海根. 1991. 生存分析理论及其在研究生命表中的应用. 生态学报, 11(2): 153-158. DOI:10.3321/j.issn:1001-9332.1991.02.013 |
Chi H, Lin H. 1985. Two new methods for the study of insect population ecology. Acad.Sin, Bull.Inst.Zool, 24(2): 225-40. |
Chi H. 1988. Life table analysis incorporating both sexes and variable development rate among individuals. Environ.Entomd, 17(1): 26-34. DOI:10.1093/ee/17.1.26 |