 2001, Vol. 37
2001, Vol. 37文章信息
- 张鼎华, 翟明普, 林平, 贾黎明, 孙志蓉.
- Zhang Dinghua, Zhai Mingpu, Lin Ping, Jia Liming, Sun Zhirong.
- 杨树刺槐混交林下沙质土壤腐殖物质特性
- HUMIC SUBSTANCES CHARACTERISTICS OF THE MIXED PLANTATION OF POPLAR AND BLACK LOCUST IN SANDY SOIL
- 林业科学, 2001, 37(3): 58-63.
- Scientia Silvae Sinicae, 2001, 37(3): 58-63.
-
文章历史
- 收稿日期:2000-06-12
-
作者相关文章


2. 新疆塔里木农垦大学植物科技学院 塔里木 843300
2. Plant Science and Technology College of Talimu Agri-reclamation University Talimu 843300
杨树(Populus spp.)刺槐(Robinia pseudoacacia L.)混交林是我国北方沙荒地造林实践中形成的一种成功的固氮树种和非固氮树种混交林, 其促进杨树生长、改善林地小气候、提高树木抗病抗虫能力、改善林地养分和水分状况以及林地景观特征等多方面作用已广为人知。为了充分认识这种混交林类型成功的奥秘以便更好地指导造林实践, 人们从它的生长状况、生物量、根系分布、土壤肥力及养分循环、小气候变化及生物化学作用方式等方面进行了一系列的研究(翟明普, 1997; 贾黎明, 1997; 金幼菊, 1997)。然而, 到目前为止, 尚未见到对该混交林沙地土壤腐殖物质特性研究的报道。本文旨在通过研究杨树刺槐混交林沙地土壤腐殖质变化状况, 了解其对土壤肥力的作用及其作用的机理, 为我国干旱、半干旱地区沙地土壤更好地培育林木提供有关土壤培肥方面的理论依据。
1 试验地概况试验地位于北京市顺义县潮白河林场, 东经116°4′, 北纬40°16′, 属暖温带大陆性气候, 四季分明, 年均气温11℃, 年降水500~700 mm, 主要集中在7~8月份。海拔20~30 m, 土壤主要为冲积沙土, 多为“通体沙”, 有机质含量低, 蓄水、保水能力差。林场范围内成林林分大多为杨树刺槐混交林。林下地被物稀少, 覆盖度2%左右, 主要有猪毛菜、灰菜、狗尾草以及禾本科植物。杨树、刺槐纯林及杨树刺槐混交林造林时间均为1980年, 营造的杨树树种均为小美旱(Populus ‘Popularis’ Chon-lin)。
2 研究方法选择土壤起源相同、地形一致、相毗邻的杨树刺槐混交林、杨树纯林、刺槐纯林以及空旷地(无林地)设置标准地, 标准地面积20 m×20 m在标准地内按等距离分层采取0~20 cm和20~40 cm土层土样, 采取时随机采取6点土样, 然后混合均匀, 每标准地重复两次。土样带回实验室后风干, 各土样土壤理化基本性质见表 1。分析时, 土壤有机质:硫酸—重铬酸钾法(中国科学院南京土壤研究所, 1978); 土壤腐殖质组成:焦磷酸钠提取—重铬酸钾法(中国科学院南京土壤研究所, 1978; 文启孝等, 1984); E4为波长465 nm处的消光系数值; Ec=E430/C, E430为430 nm波长时的消光系数, C为碳浓度(mg/mL); E4/E6=E465/E665 (为波长465 nm和665 nm处光密度值的商); RF为腐殖酸溶液在波长400 nm处的消光系数值除以30 mL该溶液所消耗的0.1mol KmnO4的mL数, ΔlogK=K400-K600, K400和K600分别为波长400 nm和600 nm处的消光系数值(Kumada, 1967) (相当于Simon的TF和RF); 土壤腐殖质结合形态:按傅积平所介绍的改进方法进行(傅积平, 1983)。
|
|
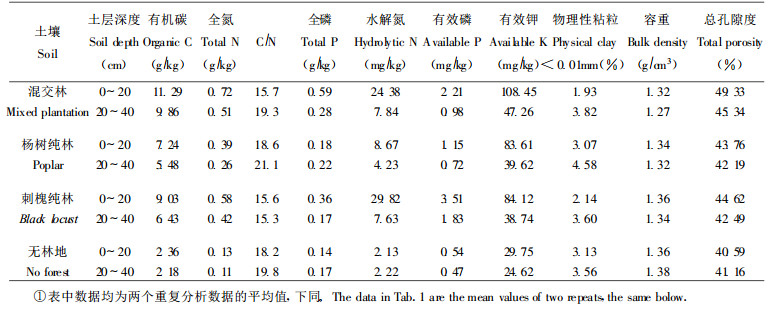
土壤腐殖质是土壤有机质的重要组成部分, 对其组成进行分析研究, 对进一步分析土壤枯落物(包括地上和地下部分)的分解状况, 揭示不同枯落物分解产物对土壤肥力的影响有一定的指导意义(文启孝, 1984)。表 2的结果表明, 沙地土壤种植杨树纯林、刺槐纯林以及杨树刺槐混交林后, 土壤有机C含量和腐殖酸C含量均有了较大幅度的增长, 与无林地相比, 混交林土壤增幅最大, 有机C和腐殖质C 0~20 cm土壤分别增加了378.4%和403.3%, 20~40 cm土壤分别增加了352.3%和377.6%;其次为刺槐纯林, 0~20 cm土壤分别增加了282.6%和303.3%, 20~40 cm土壤分别增加了195.0%和187.1%;增幅最小的为杨树纯林, 0~20 cm土壤分别增加了206.8%和213.2%, 20~40 cm土壤分别增加了151.4%和155.3%。土壤胡敏酸是土壤腐殖质的重要组成部分, 其含量及特性在一定程度上能反映土壤腐殖质的类型及性质, 土壤腐殖化度(土壤胡敏酸占土壤全碳量的百分比)则是衡量腐殖质品质优劣的标志之一(林明海, 1982)。从表 2看, 无论是胡敏酸、富啡酸还是土壤腐殖化度都以混交林为最大, 其大小顺序为:混交林 > 刺槐纯林 > 杨树纯林 > 无林地。混交林0~20 cm土壤胡敏酸含量和土壤腐殖化度分别是无林地的8.80和1.84倍, 20~40 cm土壤分别是无林地的7.65和1.69倍; 杨树纯林0~20 cm土壤胡敏酸含量和土壤腐殖化度分别是无林地的3.70和1.21倍, 20~40 cm土壤分别是无林地的3.18和1.26倍; 刺槐纯林0~20cm土壤胡敏酸含量和土壤腐殖化度分别是无林地的5.55和1.45倍, 20~40 cm土壤分别是无林地的3.88和1.32倍。土壤胡敏酸与富啡酸比值(HA/FA)的大小顺序为:混交林 > 刺槐纯林 > 杨树纯林 > 无林地, 土壤胡敏酸与富啡酸比值能在一定程度上反映土壤腐殖物质的聚合程度(熊毅)等, 1990;杨玉盛等, 1999), 土壤胡敏酸与富啡酸比值越大, 土壤腐殖物质的聚合程度也越高, 质量也越好。
|
|

上述结果表明: (1)森林植被在土壤培肥方面的作用; (2)不同的树种对土壤肥力所起的作用不同, 刺槐为豆科植物, 其培肥作用大于非豆科植物的杨树; (3)混交林对土壤的培肥作用大于纯林。混交林由于群落结构趋于复杂, 林内土壤枯落物在数量和质量上均发生变化, 导致土壤微生物数量的增加以及群体更加多样化, 因而能更加深刻地改造土壤, 提高土壤肥力(贾黎明, 1998)。
3.2 土壤腐殖质光学特性胡敏酸的光学特性是判断土壤腐殖质性质的重要依据。腐殖物质的颜色是由各种发色官能团所引起, 例如不饱和键、羰基和醌基等。胡敏酸度的光密度值在一定程度上可以表征分子的复杂程度, 胡敏酸的光密度愈大, 则分子的复杂程度愈高, 芳香核原子团多, 缩合度较高; 相反, 较为简单的胡敏酸则芳香性小, 脂肪侧键多, 光密度值小。从图 1、图 2可看出, 沙地土壤种植杨树、刺槐纯林以及杨树刺槐混交林后, 土壤胡敏酸的光密度值得以提高, 提高的幅度以混交林最大, 其次为刺槐纯林和杨树纯林。据研究(加藤秀正, 1980), “成熟”的胡敏酸光密度大, 土壤肥力持久而稳定; 而分子量较小的“幼龄”胡敏酸光密度小, 土壤肥力和熟化度低。


土壤胡敏酸的E4值可在一定程度上反映腐殖物质的芳化程度和芳化分子的大小, E4值越大, 表明腐殖物质分子的复杂程度愈高, 芳化程度大, 相反, E4值越小, 表明腐殖物质芳化程度愈小, 分子的复杂程度小(熊毅, 1990; 彭福泉, 1985)。从表 3看, 无论是0~20 cm土层还是20~40 cm土层, 土壤胡敏酸的E4值的大小顺序依次为:混交林 > 刺槐纯林 > 杨树纯林 > 无林地。与无林地相比, 混交林0~20 cm土层胡敏酸E4值增加了118.5%, 20~40 cm土层增加了113.3%;刺槐纯林0~20 cm土层胡敏酸E4值增加了66.2%, 20~40 cm土层增加了51.7%;杨树纯林0~20 cm土层胡敏酸E4值增加了33.8%, 20~40 cm土层增加了26.7%。
我国一般以土壤胡敏酸的E4/E6值作为表征土壤腐殖物质的芳构化程度的指标。而波诺马廖娃等人(波诺马廖娃, 1987; 李卫东, 1996)则认为, 把EC值作为表征芳构化的指标更为适宜, 我们对沙地土壤腐殖物质的研究支持了这一观点。从表 3可看出, 沙地各土壤腐殖物质的E4/E6值之间虽然也呈规律性的变化(混交林 < 刺槐纯林 < 杨树纯林 < 无林地), 但总的来说差异不大, 而EC值的差异却较为明显。与无林地相比, 混交林0~20 cm土层胡敏酸EC值提高了106.4%, 20~40 cm土层提高了76.7%;刺槐纯林0~20 cm土层胡敏酸EC值提高了72.4%, 20~40 cm土层提高了52.8%;杨树纯林0~20 cm土层胡敏酸EC值提高了40.2%, 20~40 cm土层提高了28.9%。EC值的大小顺序为:混交林 > 刺槐纯林 > 杨树纯林 > 无林地, 表明沙地土壤种植杨树、刺槐纯林以及杨树刺槐混交林后土壤腐殖物质的芳构化程度提高, 提高的幅度以混交林最大。
一般认为暗色物质是腐殖物质最重要的特征之一, 腐殖物质形成的本质是一种颜色逐渐变暗的过程(窦森, 1995)。Kumada等(1967)根据两项与颜色有关的指标即色调系数(ΔlogK)和相对色度(RF)将胡敏酸分为A、B、P和Rp 4个类型。从Rp到B, 或从P到A, 或从B到A, 胡敏酸的腐殖化程度加深, C/H比值提高, 分子结构趋向于复杂和成熟(熊田恭一, 1984)。从表 3可知, 按Kumada等对胡敏酸的划分, 沙地土壤无林地、刺槐纯林、杨树纯林以及杨树刺槐混交林的20~40 cm土层的胡敏酸均为P型, 杨树刺槐混交林0~20 cm土层胡敏酸的类型转变为A型。沙地土壤种植杨树、刺槐纯林后虽未能使土壤胡敏酸类型发生转变, 但与无林地相比, 色调系数(ΔlogK)的降低和相对色度(RF)的升高却是十分明显的, 表明土壤腐殖化程度加深, 胡敏酸分子变得更为复杂, 土壤的熟化度加深, 肥力提高。色调系数(ΔlogK)降低和相对色度(RF)升高的幅度以混交林土壤最大, 其0~20 cm土层胡敏酸的类型甚至转变为A型, 其次为刺槐纯林, 再次为杨树纯林。
|
|

以上土壤腐殖质光学特性的分析表明, 沙地土壤种植林木后, 土壤腐殖质分子的复杂程度提高, 分子量增大, 芳香核原子团增多, 缩合度变大, 腐殖物质的芳构化和芳化分子的复杂程度以及腐殖化程度均得到不同程度的提高, 腐殖物质的品质变佳, 土壤熟化度增大, 土壤肥力得以提高。
3.3 土壤腐殖质结合形态土壤腐殖质与无机矿物结合而形成的有机无机复合体是构成土壤肥力的基础, 是植物生长所需养分、水分来源的载体, 是土壤肥力的核心, 在土壤保肥供肥过程中起着重要的作用。不同腐殖质结合形态是有机胶体(腐殖质)与矿质胶体(粘粒)相互作用的形式, 其反映了土壤有机无机复合体中腐殖质与粘土矿物结合的松紧程度的大小。一般来说, 随着土壤肥力的提高、熟化度的增强, 松结合态的腐殖质增多, 松/紧(Ⅰ/Ⅲ)比值增大(刘树庆等, 1989)。分析结果表明(表 4), 沙地土壤的腐殖质结合形态以紧结合态为主, 松结合态为次, 稳结合态最少。从松结合态看, 无论0~20 cm土层还是20~40 cm土层, 含C量和占土壤全C的比例, 都以混交林土壤最大, 刺槐纯林为次, 杨树纯林再次, 无林地最小; 紧结合态腐殖质的含C量和占土壤全C的比例在各土壤层次中也是混交林 > 刺槐纯林 > 杨杨纯林 > 无林地, 但各土壤紧结合态腐殖质的含C量占土壤全C的比例相互之间差异不大; 稳结合态腐殖质的含C量在不同土壤层次、不同沙地土壤之间(指杨树刺槐混交林土壤、杨树纯林土壤、刺槐纯林土壤和无林地土壤之间)看不出规律性的变化, 但不同土壤层次稳结合态腐殖质的含C量占土壤全C的比例却呈现出规律性的变化, 即:无林地 > 杨树纯林 > 刺槐纯林 > 混交林; 松/紧(Ⅰ/Ⅲ)比值不同土壤层次均以混交林 > 刺槐纯林 > 杨树纯林 > 无林地。一般认为, 土壤腐殖质中松结合态的C主要来源于新鲜的有机物质, 易为土壤微生物所分解, 活性较大, 对土壤有效养分的供应起着重要的作用。土壤腐殖质紧结合态是与土壤矿物(主要是粘土矿物)部分结合较紧且较稳定的腐殖质部分, 其不易被微生物所分解, 但与土壤全量养分, 如全氮、全磷等关系密切, 故土壤紧结合态腐殖质在土壤养分的保贮及稳定土壤结构方面起着重要的作用(杨玉盛, 1999; 刘树庆, 1989)。沙地土壤粘粒含量少, 土壤的缓冲性能和保肥能力低下, 提高土壤的保肥性能是提高沙地土壤肥力的关键。以上的分析结果表明了沙地土壤种植杨树、刺槐纯林以及杨树刺槐混交林后, 土壤供肥、保肥的能力得以提高, 提高的程度以混交林最大, 其次为刺槐纯林, 再次为杨树纯林。
|
|

(1) 沙地土壤种植杨树、刺槐纯林和杨树刺槐混交林后, 土壤有机C和腐殖质C含量增加, 土壤肥力提高, 增加和提高的幅度均以混交林最大, 刺槐纯林次之, 杨树纯林最小。
(2) 沙地土壤种植杨树、刺槐纯林和杨树刺槐混交林后, 土壤胡敏酸含量、土壤腐殖化程度以及土壤胡敏酸与富啡酸比值(HA/FA)均增加, 土壤腐殖物质的聚合程度提高, 土壤腐殖物质的品质得以改善, 增加、提高和改善的幅度均以混交林最大, 刺槐纯林次之, 杨树纯林最小。
(3) 沙地土壤种植杨树、刺槐纯林和杨树刺槐混交林后, 土壤胡敏酸光密度值增加, E4和EC值提高, 色调系数(ΔlogK)降低和相对色度(RF)升高, 表明土壤腐殖化程度加深, 胡敏酸分子变得更为复杂, 土壤的熟化度提高, 增加和提高的幅度均以混交林最大, 刺槐纯林次之, 杨树纯林最小。
(4) 沙地土壤种植杨树、刺槐纯林和杨树刺槐混交林后, 土壤松结合态腐殖物质(Ⅰ)和紧结合态腐殖物质(Ⅲ)以及松/紧(Ⅰ/Ⅲ)比值均有不同程度的增加, 土壤稳结合态腐殖质的含C量占土壤全C的比例有不同程度的下降, 表明土壤供肥、保肥的能力得以提高, 提高的幅度以混交林最大, 刺槐纯林次之, 杨树纯林最小。
(5) 不同的植被群落对沙地土壤的培肥效果不同。建议今后在沙地土壤营造人工群落时, 尽可能进行多树种混交造林以及多营造一些改土培肥效果好的植物种类, 如豆科植物等, 以加速土壤肥力的提高, 改善沙地地区的生态环境。
[苏]波诺马廖娃B B, 帕洛特尼柯娃T A(魏开湄译).腐殖质与土壤形成.北京: 农业出版社, 1987
|
窦森, 陈恩凤, 须湘成, 等. 1995. 施用有机肥料对土壤胡敏酸结构特征的影响-胡敏酸的光学性质. 土壤学报, 32(1): 41-48. DOI:10.3321/j.issn:0564-3929.1995.01.004 |
傅积平. 1983. 土壤结合态腐殖质分组测定. 土壤通报, 24(2): 36-37. |
贾黎明, 翟明普, 智信等.杨树刺槐混交林生长状况及生产力.混交林研究, 北京: 中国林业出版社, 1997: 11~17
|
贾黎明, 翟明普, 胡延杰等.杨树刺槐混交林养分状况.混交林研究, 北京: 中国林业出版社, 1997: 35~41
|
贾黎明, 方陆明, 胡延杰. 1998. 杨树刺槐混交林及其纯林枯落叶分解. 应用生态学报, 9(5): 463-467. |
加藤秀正(何群摘编).日本关于腐殖质粘土复合体的一些研究.土壤学进展, 1980(1): 38~43 http://www.cnki.com.cn/Article/CJFDTotal-TRJZ198001004.htm
|
金幼菊, 陈华君, 王沙生.不同栽培条件下杨树刺槐混栽对根系生长的影响.混交林研究, 北京: 中国林业出版社, 1997: 73~77
|
李卫东, 张明亮, 逄焕成. 1996. 中国暖温带黑粘土的腐殖质特性及其与土壤发生的关系. 土壤学报, 33(4): 433-438. |
林明海, 赖庆旺. 1982. 不同熟化度红壤及红壤性水稻土的腐殖质组成及其特性. 土壤学报, 19(3): 237-247. |
刘树庆, 杜孟庸, 周健学, 等. 1989. 不同肥力土壤有机无机复合度及腐殖质结合形态及其与肥力关系研究. 土壤通报, 30(6): 267-270. |
彭福泉, 高坤林, 车玉萍. 1985. 我国几种土壤中腐殖质性质的研究. 土壤学报, 22(1): 65-73. |
文启孝. 1984. 土壤有机质研究法. 北京: 农业出版社.
|
熊毅, 李庆逵主编.中国土壤(第二版).北京: 科学出版社, 1990
|
[日]熊田恭一(李庆荣等译).土壤有机质的化学.北京: 科学出版社, 1984
|
杨玉盛, 邱仁辉, 俞新妥. 1999. 不同栽植代数29年生杉木林土壤腐殖质及结合形态的研究. 林业科学, 35(3): 116-119. DOI:10.3321/j.issn:1001-7488.1999.03.020 |
翟明普, 贾黎明, 沈国舫.杨树刺槐混交林及树种间作用机制的研究.混交林研究, 北京: 中国林业出版社, 1997: 3~10
|
中国科学院南京土壤研究所.土壤理化分析.上海科学技术出版社, 1978
|
Kumada K, Sato O, Ohsumi Y, et al. 1967. Humus composition of mountain soil in central Japan with special reference to the distribution of P type humic acid. Soil Sci.Plant Nutr., 13(5): 151-158. DOI:10.1080/00380768.1967.10431990 |