 2001, Vol. 37
2001, Vol. 37文章信息
- 贾桂霞, 沈熙环.
- Jia Guixia, Shen Xihuan.
- 华北落叶松传粉生物学的研究
- STUDY ON POLLINATION BIOLOGY OF LARIX PRINCIPIS-RUPPRECHTII MAYR
- 林业科学, 2001, 37(3): 40-45.
- Scientia Silvae Sinicae, 2001, 37(3): 40-45.
-
文章历史
- 收稿日期:2000-01-06
-
作者相关文章


落叶松属包括树种多, 分布广, 是北半球温带和寒温带的重要用材树种, 但种子产量低一直是困扰种子经营和育种工作者的问题。国内外学者对多种裸子植物的传粉机制进行过研究, 认为不同植物具有不同的传粉机制(邢树平等, 1999; 吴琼美等, 1995), 并对结实有一定的影响。在早期研究中, Hakansson (1960)把Larix leptolepis高的败育率归结为受精失败和胚胎败育。Villar et al. (1984)认为L.leptolepis种子产量低是由于有效传粉期短及受精前花粉和胚珠相互作用等因素的影响所致。Said et al. (1991)详细研究了胚珠接受花粉的能力和花粉活力, 确定造成L.eurolepis (L.decidua与L.leptolepis的杂种)种子低产的原因是花粉量不足, 并提出提高控制授粉技术可增加杂种种子产量。在对L.decidus (Barner et al., 1960), L.leptolepis (Villar et al., 1984)及L.occidentalis (Owens et al., 1979)传粉机制的研究认为, 确定裸子植物传粉机制和授粉的最佳时机, 对提高种子产量有重要意义。为提高华北落叶松种子园种子产量和遗传品质, 对其雌雄球花的空间分布、种子园中树冠周围花粉密度变化、传粉机制及不同授粉方式的种子产量及其相互关系作了研究, 并讨论了空粒产生的原因。
1 材料和方法 1.1 种子园内定点观察(1) 观察地点为内蒙古乌兰察布盟卓资县上高台华北落叶松种子园。第1作业区(2 hm2) 1976年以2年生嫁接苗定植, 19个无性系, 5m×5m顺序错位排列, 测定时无性系树高在6m左右。具体情况见(贾桂霞等, 1994)。
(2) 选取结实正常的无性系, 分别调查不同轮枝层和不同枝龄的雌雄球花量。其中对轮枝层和枝龄上花量的分布, 调查了3个无性系, 3次重复, 共9株树。
(3) 在种子园作业区的中心部位, 选择无性系, 在其树冠周围不同方位、不同高度设置垂直和水平方向玻片, 每12 h取样1次, 取10个视野的平均值, 观测花粉密度的时空变化。
(4) 选择无性系, 每日定时观察雌雄球花的形态变化, 跟踪记录开花过程。
1.2 珠被顶端解剖观察采集传粉期不同发育阶段的雌球花, FAA固定, 分别进行扫描电镜(SEM)和光镜(LM)观察。SEM:将已固定的胚珠和珠鳞剥出, 系列酒精脱水, 临界点干燥, 喷金, 在Hitachi-S 800扫描电镜下观察。LM:石蜡包埋, 切片厚度8 μm, 高碘酸-锡夫试剂(PAS)和考马斯亮兰对染, 以显示传粉期胚珠中淀粉和蛋白质的动态变化。
1.3 选择3个无性系, 3次重复, 分别测定了自由授粉、辅助授粉和控制授粉种子饱满率切取花枝在室内水培, 分号收集花粉。对控制授粉的无性系, 于花粉飞散前1周左右去雄套袋。当雌球花进入最佳可授期时, 进行授粉, 第2天重复1次, 授粉半月后, 用细目化纤防虫罩换下花粉隔离袋。自由授粉的雌球花于开花前套好防虫罩。对辅助授粉的无性系, 用混合花粉授粉。9月中旬种子成熟时, 连同防虫罩一起采回, 分组调制种子。用X光摄影法观测与实体解剖以检测各组合种子的饱满率。
1.4 胚珠接受的花粉粒数传粉期后采集不同授粉方式、树冠周围不同方位和高度的球果, 剥出胚珠, 在珠孔处切一小口, 使珠心和花粉室露出, 滴0.5 %饱和水合氯醛酸性品红溶液, 复盖玻片, 轻压挤出花粉, 镜检。
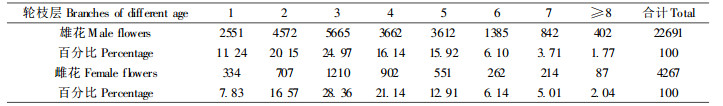
2 结果与分析 2.1 雌雄球花的分布规律华北落叶松为单性花, 雌雄同株, 风媒。雌雄球花的分布与枝龄有极显著的关系。树冠外层的1龄枝基本不着生球花, 雌球花一般着生在2~4龄枝, 所占比例为89.5%, 在5龄以上枝条上很少着生雌球花。雄球花分布的范围较雌球花广, 主要分布在2~7龄枝上, 其中2~4龄上的雄花占总雄花的82.6%。随新枝的形成, 第2年的结实层向外扩展1个年枝段。
花芽的分布不仅与枝龄密切相关, 而且与年枝层也有很大关系, 结果见表 1。雌球花主要分布在树冠中部, 且集中在2~5轮枝层; 而雄球花基本上是全冠位分布, 树冠中下部较多。随树龄的增加, 雌雄球花有向上推移的趋势。华北落叶松雌雄球花的这种分布特点, 构成了结实功能有别的树冠层次, 即以树干为轴心, 由里向外, 可划分为结实衰退区、结实区和枝条生长区, 并随年龄的增加, 3个区的大小处在动态发展中。树冠中上部的2~4龄枝是分化雌花芽最强的部位, 随枝龄的增大, 枝条分化雌花能力渐衰, 就整株树而言, 表现为雌球花开花部位纵向上移, 横向外移, 稳定在较幼龄枝条的区内, 即结实区, 而内膛结实衰退区不断扩大。雄球花分布较广, 多分布在树冠中下部的2~7龄枝上, 且随树龄增加, 有由下而上、由内向外推移的趋势, 但推移速度较雌球花缓慢。
|
|
4月底, 随着雄球花轴迅速伸长, 小孢子囊壁破裂, 进入散粉期。由于散粉期间温度、湿度和风速的不同, 在不同年份不同地点, 散粉的历程有所不同, 传播距离也不同。95%的花粉在3~4d内飞散, 整个花期延续6~7d。1d内, 大量散粉出现在8:00~18:00间, 且最大值多在10:00~12:00间。
为了解种子园中雌球花授粉的机率与花粉云密度的关系, 测定了种子园内树冠周围花粉的空间变化, 选取东、南、西、北4个方向, 分别测定距地面1、2、3、4、5m高处的花粉密度, 结果见图 1。图 1中看出, 在树冠周围3m处花粉密度最大, 5m处最小。这与雄花分布特点和花粉结构有关, 华北落叶松的花粉无气囊, 且多分布在树冠中下部, 受重力影响较大, 飞扬不高。同一高度不同方位花粉密度也有一定差异, 东、南方向的较大, 西、北方向相对小一些, 这与当地的气候条件有关, 散粉期, 多为西、北风, 从而使树冠周围花粉密度受风向影响较大, 造成东南西北4个方位各占26.8%, 30.58%, 21.88%, 20.67%。观测结果显示, 当风力较小时, 水平方向和垂直方向所接受的花粉数量差别不大, 但如受大风(6~7级)影响, 垂直方向接受的花粉量明显多于水平方向, 说明大部分花粉受风的影响倾斜下降。

|
图 1 树冠周围不同方位与不同高度花粉密度的分布 Fig. 1 The distribution of pollen density at different heights and directions around the crown

|
雌球花具大孢子叶50~60枚, 螺旋状排列, 每个大孢子叶有两个倒生的胚珠。经休眠后的胚珠, 第2年春天开始生长, 4月底、5月初进入传粉期。
据外部形态特征, 结合解剖结构和内部物质的变化, 可将进入传粉期的雌球花基本划分为以下5个阶段:
(1) 雌球花从芽鳞中露出, 只能看到苞鳞的尖端, 此时进入初花期。珠被原基发育较快, 形成珠被, 且在胚珠的顶端不均等发育, 即上面(远轴面)为唇状的小裂片, 下部(近轴面)发育为大裂片, 大裂片将成为接受花粉的主要场所, 裂缝状的珠孔道存在于两者之间, 但通常被下面的大裂片所掩盖(图版Ⅰ-1)。

|
图版Ⅰ Plate Ⅰ 1-4、7.为扫描电镜照片, 5、6为经PAS染色的光学显微照片。1.进入传粉期第1阶段的胚珠, 两侧的珠被在胚珠顶端开始不均等发育(空心箭头), ×300。2.近轴面的珠被顶端发育较快, 形成较大的裂片, 且其表面细胞突起, ×350。3.大裂片继续增大, 并形成表皮毛, ×150。4.处于最佳传粉期的胚珠顶端, 大裂片上的表皮毛充分发育, 以接受花粉, ×220。5.进入传粉第2阶段的胚珠, 示近轴面的珠被顶端的大裂片, 且细胞中含有一定量的淀粉颗粒(箭头所示), ×165。6.传粉末期的胚珠, 花粉随珠被毛的内折进入到珠孔道(箭头), 珠被毛萎缩退化, 形成粘性物质, 与PAS反应强烈(空心箭头), ×165。7.示传粉末期的胚珠顶端, 珠被毛内折, ×170。 The pollination process of Larix principis-rupprechtii Mayr. Plate 1-4, 7 are scanning electron micrographs and plate 5, 6 are light micrographs stained with PAS, showing the change of starch grain during pollination. 1.The ovule with unevenly developing integuments, a larger one (hollow arrow) and a smaller one, which was at first stage of pollination, ×300.2.Extension of larger integument to form lobe at its upper part with papillae, ×350.3.Increment of lobe with the elongation of palippae, ×150.4.The top part of ovule at optimal time of pollination, showing fully developed papillae on the surface of lobe to receive pollens, ×220.5.Starch grains in the cells of larger integument lobe, showing the ovule was at second stage of pollination, ×165.6.Pollen brought into the micropylar canal (arrow)with the engulfment of papillae at the end of pillination.Papillae degenerated to form sticky materials which reacted with PAS strongly (hollow arrow), ×165.7.Morphology of top ovule at the end of pollination, showing engulfment of papillae, ×170. |
(2) 雌球花向上弯曲生长, 节间伸长, 可看到种鳞和球花的底部。大裂片继续增大, 其顶端表皮细胞突起, 形成短的珠被毛, 经PAS反应, 显示在大裂片中具有较多的淀粉颗粒和多糖物质(图版Ⅰ-2、5)。此时种鳞螺旋状排列变得更明显, 使靠近球花轴的珠被顶端同样成为螺旋状排列, 增加了花粉落入的空间。雌球花基部和顶端的节间短, 胚珠大多发育欠佳, 只有退化的胚珠, 而无珠被毛。
(3) 雌球花直立, 节间继续伸长, 种鳞之间彼此分开, 胚珠露出, 手感较柔软。珠被顶端的大裂片变为较大的球状, 在大裂片和珠被本体过渡区部分形成颈的结构, 珠被毛伸长, 是发育最盛的时期。在裂片和珠被毛中仍有少量的淀粉, 且珠被毛表皮上分泌有多糖类物质(图版Ⅰ-3、4)。随球花轴节间伸长, 种鳞之间有空间增大, 使花粉进入, 落入的部分花粉被珠被毛所粘附。这一阶段为最佳传粉期, 据观察, 一般持续2d左右。
(4) 雌球花直立, 种鳞增厚, 种鳞之间的距离减小, 雌球花开始变硬。大裂片基部的表皮细胞伸长并向珠孔道弯曲, 这使得珠被毛及其粘附的花粉开始内折, 进入到珠孔道(图版Ⅰ-7)。此时, 大裂片和珠被毛中的淀粉消失, 但仍有大量的多糖物质。部分与花粉接触的珠被毛退化萎缩, 形成一堆粘性物质。经PAS反应显示, 为大量的糖类物质(图版Ⅰ-6)。
(5) 种鳞继续增厚, 封闭彼此间的空隙, 传粉结束。珠被顶端的珠被毛继续内折, 直到整个珠被顶端进入珠孔道, 粘附在珠被毛上的花粉一同进入到珠孔道, 此时珠孔道关闭。
从1~5阶段, 约持续1周, 在整个传粉过程中, 未观察到传粉滴从珠孔道分泌出来, 这与多数裸子植物的传粉机制不同, 它主要依靠珠被大裂片顶端突起的表皮毛及其分泌的粘性物质来捕获花粉。最佳传粉期在第3阶段, 约持续2d。
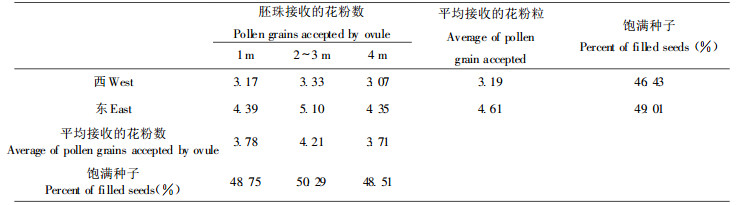
2.4 胚珠授粉状况与种子产量解剖5月底着生在树冠不同方向和高度的胚珠, 检查其中所接受的花粉粒数, 并在同年8月底球果成熟时, 采集相应位置的球果作分析, 结果见表 2。从表中可以得出, 种子园中雌球花捕获的花粉数量在不同方向和不同高度均有所不同, 这一结果与不同空间花粉密度的差异是相对应的, 从而也说明了种子园中雌球花授粉机率与花粉密度有一定的关系。
|
|
为了解同一个球果内不同部位的种子生产情况, 对球果上、中、下3部分别检测了胚珠接受的花粉量, 具体接受情况见图 2。

|
图 2 球果不同部位胚珠接受花粉的频度 Fig. 2 The frequency of ovules accepted pollen grains at different parts of cone(Grain)

|
从每个胚珠接受的花粉数看, 球果中部平均为5粒左右, 多数变动于3~7粒, 占73.3%, 没接受到花粉的胚珠为3%;上、下两部分平均是3~4个, 上部0~5个占82.7%, 其中0个占10.7%, 基部变动于0~6个, 其中0个为11.2%。
同年当种子成熟时, 采集相应部位的球果, 同样分上、中、下3部, 检测种子的类别, 结果见表 3。
|
|

球果上、下两端由于不育种鳞的存在, 胚珠发育不完全, 上、下部位各有10.7%和11.2%的胚珠接受不到花粉, 造成残翅和胚珠败育, 这和表 3中所列的上、下部残翅和败育百分率比较一致。此外, 上、中、下3部均有空粒存在, 差异不大, 变动于9.33%~12.91%, 其中中部所占比例小, 为9.33%, 上部较大, 占12.91%, 下部为10.41%, 由此得出, 胚珠接受的花粉数与种子产量有直接的关系, 但并不是造成空粒的主要原因。
3 讨论与结论据观察, 上高台华北落叶松种子园中, 最佳可授期在2 d左右, 这样短的可授期是否影响花粉的接受, 从检测结果看, 胚珠平均接收的花粉数3~4粒, 中部未接收到花粉的胚珠平均为3%, 上下两部分由于不育种鳞的存在, 分别为10.7%和11.2%, 表明当种子园进入结实盛期, 花粉量充足时, 2 d可授期内多数胚珠能接收到足够的花粉; 但当花粉量不足时, 可能会成为主要问题。在Owens et al. (1993)的研究中, L.occidential珠果的可授期仅为3~5 d, 最佳可授期接近3d;Villar et al. (1984)提出在L.leptolepis种子园中, 结实率低是由于可授期短, 他们认为, 在该试验条件下, 可授期少于1 d, 辅助授粉就成为提高种子产量的重要手段。
虽然辅助授粉是提高种子园种子产量和质量的重要手段, 但授粉时机的掌握至关重要。对那些具有传粉滴的裸子植物, 可根据传粉滴的出现来确定可授期。然而华北落叶松在传粉期并不产生传粉滴, 因此必须依据雌球花的形态变化和胚珠珠被顶端的变化来准确地确定最佳可授期, 否则收效不大。试验中分别测定了自由授粉、辅助授粉和控制授粉的授粉效果及种子生产情况, 结果见表 4。套袋控制授粉, 虽然胚珠平均接收的花粉粒数较多, 有6个, 但由于授粉时机掌握不好, 未捕捉到花粉的胚珠数量相当大, 占21.1%, 再加上套袋使袋内环境不同于外部, 影响种子早期发育, 造成饱满种子率很低。因此, 对控制授粉, 应多次授粉, 并提高授粉技术。
|
|

以上结果表明, 确定最佳可授期很重要。从形态上看, 第3阶段的雌球花苞鳞之间彼此分开, 可看到球花轴和倒生的胚珠, 苞鳞近似水平, 与球花轴基本成直角。用手持放大镜就可观察到胚珠珠孔顶端的变化, 当珠被毛伸长且具有粘液时, 即到达可授期。另外球花发育的时期与温度有很大关系, 授粉时还应考虑自然条件和套袋内外温度的差, Runions et al. (1996)在对云杉传粉机制的研究中, 观测到袋内雌花最佳可授期早于自然条件的雌花2~3d。
上述研究说明, 饱满种子的产量与胚珠接受的花粉量有关, 却不是造成种子空粒的主要因素。要弄清产生空粒的原因及其机制, 仍需从其它方面做研究。
贾桂霞, 沈熙环, 李凤兰. 1994. 华北落叶松雌雄配子体的形成及胚胎发育. 北京林业大学学报, 16(2): 10-14. DOI:10.3321/j.issn:1000-1522.1994.02.001 |
邢树平, 张泉, 胡玉熹, 等. 1999. 侧柏和北美香柏(柏科)的传粉机制. 植物学报, 40(2): 130-132. DOI:10.3321/j.issn:1672-9072.1999.02.003 |
吴琼美, 樊汝汶, 肖石海, 等. 1995. 杉木传粉生物学的观察. 南京林业大学学报, 19(1): 47-51. |
Barner H, Christiansen H. 1960. The formation of pollen, the pollination mechanism and the determination of the most favorable time for controlled pollination in Larix. Silvae Genet., 9: 1-11. |
Hakansson A. 1960. Seed development in Larix. Bot.Not., 113: 29-40. |
Owens J N, Molder M. 1979. Sexual reproduction of Larix occidentalis. Can.J.Bot, 57: 2673-2690. DOI:10.1139/b79-317 |
Owens J N, Morris Sh. J, Catalano G L. 1993. How the pollination mechanism and prezygotic and postzygotic events affect seed production in Larix occidentalis. Can.J.For.Res., 24: 917-927. |
Runions C J, Owens J N. 1996. Pollen scavenging and rain involvement in the pollination mechanism of interion spruce. Can.J.Bot., 74: 115-124. DOI:10.1139/b96-016 |
Said C, Villar M, Zandonella P. 1991. Ovule receptivity and pollen viability in Japanese larch (Larix leptolepis Gord). Silvae Genet., 40: 1-6. |
Villar M, Knox R B, Dumas C. 1984. Effective pollination period and nature of pollen-collecting apparatus in the Gymnosperm, Larix leptolepis. Ann.Bot.(London), 53: 279-284. DOI:10.1093/oxfordjournals.aob.a086689 |