 2001, Vol. 37
2001, Vol. 37文章信息
- 徐庆, 刘世荣, 臧润国, 郭泉水, 郝玉光.
- Xu Qing, Liu Shirong, Zang Runguo, Guo Quanshui, Hao Yuguang.
- 中国特有植物四合木种群的生殖生态特征———种群生殖值及生殖分配研究
- THE CHARACTERISTICS OF REPRODUCTIVE ECOLOGY OF ENDEMIC SPECIES TETRAENA MONGOLICA POPULATION IN CHINA———Reproductive Value and Reproductive Allocation
- 林业科学, 2001, 37(2): 29-35.
- Scientia Silvae Sinicae, 2001, 37(2): 29-35.
-
文章历史
- 收稿日期:2000-05-09
-
作者相关文章


2. 中国林业科学研究院沙漠林业实验中心 磴口 015200
2. Experimental Center of Desert Forestry of CAF Dengkou 015200
四合木(Tetraena mongolica Maxim.)是蒺藜科(Zygophyllaceae)中单种属植物, 是我国内蒙古所特有的国家二级保护植物。主要分布在鄂尔多斯高原西北部, 库布齐沙漠以南, 桌子山(阿拉巴素山)的山麓地带, 少量延伸到相邻的乌达低山残丘区。在草原化荒漠地区, 可形成以四合木为建群种的植物群落。
植物种群的更新和种群中植物个体生活史的完成依赖于生殖过程。在生殖过程中的生殖分配格局以及植物如何调节其生殖分配以适应特定生存环境, 是生殖生态学研究的重要任务(苏智先, 1998)。生殖分配(Reproductive Allocation) :指植物在生长发育过程中, 同化产物向其生殖器官分配的比例, 即分配到生殖器官中的有机物数量。它控制着植物终生生殖与生存的平衡(江洪, 1992)。生殖值(Reproductive Value)是指一个平均年龄的个体, 在其死亡之前对下一代的平均贡献(Silvertown, 1982), 或指某年龄的成员从现在到死亡, 对下一世代的贡献值(江洪, 1992;臧润国, 1993)。生殖值的计算, 是研究种群生殖的重要手段, 它不仅可以计算现实的生殖能力, 而且还可以对种群的潜在生殖能力做出推测。
生殖值、生殖分配的研究是生殖生态学中的热门领域, 在草本植物的生殖值、生殖分配方面成果更加突出, 但对乔、灌木的研究甚少(苏智先, 1998), 尤其对四合木种群生殖分配、生殖值的研究, 目前尚无报道。本文试图从生殖生态学的角度, 对四合木种群生殖值、生殖分配与生殖阶段、生殖年龄等方面进行研究, 以探讨四合木种群最适生殖格局及生殖对策, 以期为保护和利用该物种提供理论依据。
1 研究地区概况研究地区位于内蒙古杭锦旗丘地及滩地, 地理位置为北纬39°45′~40°20′, 东经106°40′~107°20′。本区大陆性气候明显, 冬季寒冷, 夏季酷热, 干旱少雨, 风大沙多, 热量丰富。全年日照总时数平均为2792.2 h, 年平均温度9.7 ℃, 极端最高气温达39 ℃, 极端最低气温达-32 ℃; 地面最高温度达63.4 ℃, 地面最低温度为-23.4 ℃, 年平均降水量137.8 mm, 大都集中在6~8月; 年均蒸发量3217.7 mm, 干燥度4.05。基岩为砂岩, 土壤以灰漠土为主。
杭锦旗丘地的群落(Ⅰ)特点为:四合木为优势种, 上层有红沙(Reaumuria soongorica)、狭叶锦鸡儿(Caragana stenophylla), 其下有珍珠柴(Salsola passerina)、骆绒藜(Ceratoides latens)伴生。多年生丛生禾草形成亚优势层片, 主要代表有无芒隐子草(Cleistogenes songarica)、沙生针茅(Stipa glareosa)、戈壁针茅(Stipa gobica)、短花针茅(Stipa breviflora)等。
杭锦旗滩地的群落(Ⅱ)特点为:霸王柴(Zygophyllum xanthoxylon)、四合木为优势种, 并与少量沙冬青(Ammopiptanthus mongolicus)构成群落的上层; 其下有绵刺(Potaninia mongolica)、刺叶柄棘豆(Oxytropis aciphy)等小灌木; 无芒隐子草、沙生针茅、戈壁针茅等多年生禾草在下层形成亚优势层片; 其它草本成分有戈壁天门冬(Asparagus gobicus)、黄蒿(Artemisia scoparia)、草霸王(Zygophyllum mucronatum)、阿氏旋花(Convolvulus ammannii)、蒙古葱(Allium monglicum)和多根葱(Allium polyrrhizum)等。调查样地的群落及生境特征见表 1。
|
|
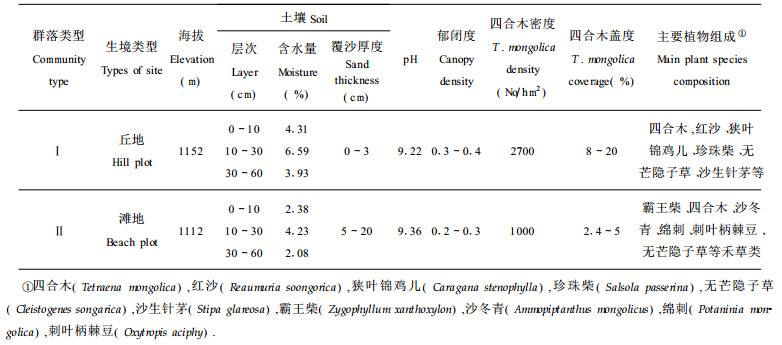
根据群落调查方法, 在丘地及滩地各设置10 m宽样带1个, 沿样带每隔100 m设置10 m×10 m的样方1个。每生境样方各35个, 精确测定各样方每株四合木的地径、冠幅, 其中地径用游标卡尺测量, 高度及冠幅用钢卷尺测量, 同时调查其群落植物组成, 统计四合木的株数, 每生境沿样带对角线设5个采集点, 按0~10 cm, 10~30 cm和30~60 cm 3个层次分别取样, 用称重法测定土壤含水量; 土壤:水(1:2.5)制成泥浆后, 用pH计测定其pH, 同时测定土壤表层覆沙厚度、海拔等生境指标。
截取77个四合木标准植株基径样品, 做好标记, 带回实验室进行切面、磨光, 用瑞典产的年轮分析仪(CXDDO型)查数年轮, 建立年龄(A)与地径(D)的回归方程:A=0.14+0.6709D, (R=0.91886, n=77, p < 0.05), 以此方程为基础, 计算所调查地区每株四合木的年龄, 以3a为1龄级对四合木种群进行龄级划分, 统计各龄级株数及x年龄开始时标准化的存活率Lx。
在丘地及滩地中每个龄级选择标准植株5株以上, 测其地径, 在果实未脱落之前, 分别采集测定果实、种子数, 统计各龄级平均产种子数(Mx), 测定其年净生长量, 即把当年生长枝、叶、花或果产量进行统计(本研究将当年生新枝产量、叶、生殖器官部分分别进行统计), 根据年轮确定茎当年生长量, 分别称鲜重后, 在烘箱内105℃烘至恒重, 用日本产“KS-016A型”电子天平称其干重。
生殖值(Vx)可用下列公式来计算(江洪, 1992;臧润国, 1993) :
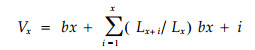
|
式中, Lx+i/Lx为年龄x到x+i的生存概率。在稳定的种群中, 年龄为0的个体生殖值与净生殖率R0相等, 繁殖终止时, 残有的个体生殖值为0。bx+i和bx分别表示年龄x+i和x时, 种群的平均生殖力。即生殖力表中的Mx+i和Mx, 这些统计数据可从种群的生命表和生殖力表中获得。Vx为现时年龄期内所产生子代的平均数bx+i之和, 同时把年龄x的个体将存活到那些年龄期的每一个年龄期的概率(Lx+i/Lx)计算在内。生殖值可分解为现时生殖值(Present reproductive value)与剩余生殖值(Residual reproductive value)。前者可记为M, 表示当年产生的子代数, 实际上就是生殖力表中的Mx, 后者可记为RRV, 表示以后所有生殖季节产生的子代数(Silvertown, 1982; Willson, 1983)。Vx=M+RRV。
植物对资源的分配原则表明, 环境资源主要用于植物3种活动:维持生命(maintenance of life)、生长(growth)和生殖(reproduction)。如果总资源量相对恒定, 对生物生殖所必需的资源配置量的增加, 必然导致其它两个过程(即维持和生长)资源配置量的减少, 其中用于维持生命的资源部分是固定的(苏智先, 1998)。生殖分配(RA)一般用下式进行计算:

|
经我们在研究区域的大量调查, 发现四合木初次结实年龄为4a, 3a以下的四合木无开花结实者, 以3a为1龄级, 根据所调查的各龄级四合木的株数及平均每株四合木产种子的数量(现时生殖值或生殖力Mx), 统计出杭锦旗丘地及滩地四合木种群生殖值(如表 2), 其中包含各龄级四合木存活的概率(Lx)、生殖值(Vx)与剩余生殖值(RRV)。
|
|
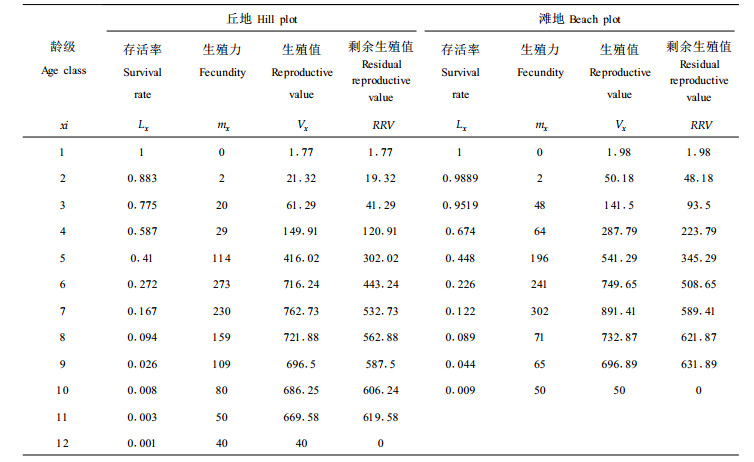
从表 2可以看出, 1龄级时, 因3a以下植株未开花结实, M为0, Vx与RRV几乎接近于0, 但由于有了一定的值, 说明已有生殖的可能性, 但在现有的条件下没有变成现实性。2龄级以后, Vx与RRV值都较大, 在现有条件下, 原来的可能性已经变成现实性, 所以Vx也较大。7龄级时, Vx值为最高, 丘地Vx为762.73, 这一数据可以比龄级为1时的Vx高出431倍。滩地Vx为891.41, 比龄级为1时的Vx高出450倍。但到达这一生殖值高峰后, 便出现缓慢下降的趋势, 到10龄级时, 滩地Vx为50, 即等于现时生殖值(Mx), 该群落中再没有更大年龄的四合木植株, 所以, RRV剩余生殖值为0。丘地到12龄级时, Vx为40, 等于现时生殖值(Mx), 该群落中再没有更大年龄的四合木植株, 所以, RRV剩余生殖值为0。如果以M≠0, 且Vx值大于Vx平均值的龄级算生殖高峰期, 那么丘地Vx平均值411.96, 生殖高峰期为5~11龄级(13~33a); 滩地Vx平均值为414.36, 生殖高峰期为5~9龄级(13~27a)。
以Vx为纵坐标, 以龄级为横坐标, 绘制Vx随龄级的变化图, 如图 1所示。

|
图 1 四合木种群的Vx动态 Fig. 1 Dynamics of Tetraena mongolica population ---▲---滩地Beach plot; —●—丘地Hill plot. |
从图 1可以看出, 丘地、滩地2个不同群落中的四合木种群的生殖值Vx都是中间高, 两头低, 随龄级的变化曲线呈山峰型。
在相同龄级, 不同生境、不同群落类型中的四合木种群的生殖值也存在差异。杭锦旗滩地四合木种群在27 a内各龄级的生殖值比相同龄级丘地的生殖值高, 30 a内总生殖值也高于生境条件较好的丘地。这是因为: (1)在滩地, 群落中霸王柴为优势种, 有同科植物霸王柴与之竞争养分、水分, 不利于四合木种群的更新。(2)滩地土壤含水率(2.87)低于丘地土壤含水率(4.94), 加上附近乌兰布和沙漠风沙的影响, 覆沙厚度较高(5~20 cm), 持水性较丘地差。四合木种群在生境条件较差的群落中生殖值较高, 这可能是四合木为了生存所采取的生殖对策。
以丘地四合木种群生命表中的Lx、kx为纵坐标, 以龄级为横坐标, 绘制Lx、kx随龄级的变化趋势, 如图 2。从图 2可以看出, 生殖值Vx随龄级的变化趋势基本上与环境选择压力(致死力kx)随龄级的变化趋势基本一致, kx小时Vx小, kx大时Vx大; 生殖值Vx随龄级的变化与存活率Lx随龄级的变化趋势相反, Lx小时Vx大, Lx大时Vx小, 通过生命表和生殖值的分析, 我们可以看出, 四合木种群的生殖值受环境因子的选择压力(kx)以及种群自身的存活率(Lx)所控制的。

|
图 2 四合木种群的Lx、kx随龄级变化图 Fig. 2 The changes in Lx、kx with age-class of Traena mongolica population —▲—环境选择压力kxSelective pressure of environment (kx); —●—存活率LxSurvival rate. |
四合木种群进入生殖期后, 每年均开花结实(5~6月开花, 7~9月结实), 但花期和果期两个不同生殖阶段生殖分配格局的变化规律不同。
花期:丘地及滩地两个不同四合木种群在花期相同龄级3~6龄级(9~18 a)的四合木个体地上年净生物产量在不同器官(枝1、叶1、花)中的分配, 如图 3。图 3表明, 丘地:枝1为29.01%、叶1为59.98%、花为11.17%;滩地:枝1为34.51%、叶1为51.54%、花为13.64%, 从RA来看, 丘地:RA=11.17%, RA值较低, 即分配到花这一生殖器官的净生产力占地上总净生产力的11.17%。滩地:RA=13.64%, 分配到花这一生殖器官的净生产力占地上总净生产力的13.64%。果期:四合木种群果期3~6龄级(9~18 a)的个体地上年净生物产量在不同器官(枝2、叶2、果)中的分配(图 3)研究表明, 丘地:枝2为32.87%、叶2为60.75%、果为6.38%;滩地:枝2为32.05%、叶2为60.95%、果为7%, 果期地上年净生物量生殖分配格局与花期相比有变化, 生殖器官(果实)的生物量占地上总净生物量的比率减少, 杭锦旗丘地生殖分配(RA)由花期的11.17%降到果期6.38%, 滩地由花期的13.95%降到果期的7%。产生这一现象的原因可能包括: (1)果期四合木种群地上净生产力分配给营养器官的比率增加, 尤其是叶, 所以导致生殖器官生产力分配相对减少; (2)四合木是雌雄同株植物, 花果转移率在69%以下, 因而需大量的花来保证其果实及种子数量, 所以花期分配给生殖器官的比率大。(3)四合木花期叶小而少, 果期叶大而多, 果实、种子小, 重量轻(1.1121g/1000粒)。(4)花的后期, 胚珠败育现象严重, 胚珠发育成种子寥寥无几, 四合木种群地上净生产力分配给胚珠已败育的花、果就失去了意义, 不如将多余的能量分配给营养器官, 为下一年的生殖积累能量, 这也是四合木维持其生存所采取生殖对策。

|
图 3 四合木种群花期、果期的地上生物量分配 Fig. 3 The allocation of aboveground biomass of Tetraena mongolica population at flowering and fruiting phase 枝:Branch; 叶:Leaf; 花:Flower; 果:Fruit. □丘地Hill plot; ■滩地Beach plot. |
在花、果这两个不同的生殖阶段, 就丘地、滩地而言, 其生物量的分配总的趋势为:叶〉枝〉花。叶的生物量配置一直居首, 变化在51.54%~60.95%。可见叶是营养器官中最重要的功能器官, 是生殖过程中营养物质的主要供应者。在草原化荒漠地区, 干旱少雨, 四合木的叶虽小, 但肉质叶贮水功能较强, 除了保证其持续地向其它器官(尤其生殖器官)输送营养物质外, 还在一定程度上为这些器官供应部分水分。生殖器官RA值, 从花期到果期, 呈下降的趋势。
3.2.2 生殖年龄与生殖分配以丘地、滩地四合木种群果期地上净生物量为基础。计算出生殖分配值(RA), 以3年为1龄级, 绘制年龄为RA关系图(图 4)。图 4表明, 杭锦旗丘地四合木种群在4~21 a之间, 其RA值随年龄增大逐渐递增, 21 a以后, 其RA值随年龄增大逐渐递减, 在15~27 a, 其RA值占总RA值的80.34%, 即为生殖高峰期。滩地四合木种群在4~9 a及12~21 a之间, 其RA值随年龄增大逐渐递增。

|
图 4 四合木RA值随龄级动态变化 Fig. 4 The dynamic change of RA with age class for Tetraena mongolica —▲—滩地Beach plot; —●—丘地Hill plot. |
但10~12 a时出现下降波动。21 a以后, RA值随年龄增大逐渐递减, 在13~27 a的RA值占总RA值的66.52%。由此进一步表明, 杭锦旗丘地、滩地四合木种群的年龄在4~6年, 其生殖器官发育成熟, 在13 a以下时, 其RA值较低, 尤其9 a以下, 滩地RA < 5.61%;丘地RA < 2.41%;13~27a之间, RA值为7.37%~11.14%, 即生殖值相对稳定; RA>21 a时, RA值为3.02%~5.20%, 较前一个阶段下降。
根据生殖年龄和生殖分配RA值的大小, 可把四合木种群的生殖期分为生殖起动期(< 9 a), 生殖增长期(9~12 a), 生殖高峰期(13~27 a)和生殖衰退期(>27 a)。
4 结论四合种群的生殖值从低龄组到高龄组, 呈现出一种低→高→低的趋势。3~9 a, 生殖值缓慢增加, 10~21 a, 生殖值迅速增高, 19~21 a, 达到最大值, 21 a以后, 生殖值缓慢下降并渐渐平缓的趋势, 繁殖终止时残有的个体, 其生殖值为0, 生殖高峰期在13~27 a。四合木种群生殖值受环境因子的选择压力(kx)以及种群自身的存活率(Lx)所控制的。
根据其生殖年龄和生殖分配RA值的大小, 可把四合木种群的生殖期分为生殖起动期(〈9 a), 生殖增长期(9~12 a), 生殖高峰期(13~27 a)和生殖衰退期(>27 a)。
在环境恶劣的滩地, 四合木种群的生殖值、地上年净生物量的生殖分配(RA)比生境条件好丘地要高, 这可能是四合木种群为了生存其生殖过程所采取的适应对策。
曹坤方. 1993. 植物生殖生态学透视. 植物学通报, 10(2): 15-23. |
方炎明. 1996. 植物生殖生态学. 济南: 山东大学出版社.
|
江洪. 1992. 云杉种群生态学. 北京: 中国林业出版社.
|
苏智先, 张素兰, 钟章成. 1998. 植物生殖生态学研究进展. 生物学杂志, 17(1): 39-46. |
苏智先, 钟章成. 1998. 四种大头茶种群生态学研究Ⅱ. 种群生物量生殖配置格局研究, 生态学报, 18(4): 379-385. |
吴素琴, 李克昌, 杨瑞全. 1994. 四合木种子特性的测定研究. 草业科学, 11(3): 29-31. |
臧润国, 李德志, 宋树强. 1993. 天然次生林群落中刺五加种群生态学的研究(Ⅳ)—刺五加种群的生殖值及其变化. 吉林林学院学报, 9(2): 7-10. |
朱宗元, 雍世鹏, 刘钟龄.四合木荒漠Form.Tetroena mongolica内蒙古植被.内蒙古宁夏综合考察队编.北京: 科学出版社, 1985, 687~690
|
Abrahamson WG. 1982. On the comparative allocation of biomass, energy, and nutrients in plant. Ecology, 63(4): 982-991. DOI:10.2307/1937238 |
Bazzaz FA, et al. 1979. Contribution to reproductive effort by photosynthesis of flowers and fruits. Nature, 279: 554-555. DOI:10.1038/279554a0 |
Douglas DA. 1971. The balance between vegetative and sexual reproduction of Mimulus primuloides(Scrophulariceae)at different altitude in California. J.Ecol., 69: 295-310. |
Hegazy AK. 1992. Age-specific survival, mortality and reproduction, and prospects for conservation of Limonium delicalumlum. J.App.Ecol, 29: 549-557. DOI:10.2307/2404462 |
Silvertown JW, Dous JL. 1982. Introduction to plant population ecology, Blackwell scientific publication. Oxford. |
Silvertown JW. 1982. Introduction to plant population ecology. Longman, London and New York. |
Willson MF. 1983. Plant reproductive ecology.John wiley and Sons. New York. |