 2001, Vol. 37
2001, Vol. 37文章信息
- 史作民, 刘世荣, 程瑞梅, 蒋有绪.
- Shi Zuomin, Liu Shirong, Cheng Ruimei, Jiang Youxu.
- 宝天曼落叶阔叶林种间联结性研究
- INTERSPECIFIC ASSOCIATION OF PLANT POPULATIONS IN DECIDUOUS BROAD-LEAVED FOREST IN BAO TIANMAN
- 林业科学, 2001, 37(2): 20-28.
- Scientia Silvae Sinicae, 2001, 37(2): 20-28.
-
文章历史
- 收稿日期:1999-09-08
-
作者相关文章


种间联结(interspecific association)是指不同物种在空间分布上的相互关联性, 通常是由于群落生境的差异影响了物种的分布而引起的(王伯荪等, 1989; Greig, 1983)。这种联结性就是对各个物种在不同生境中相互影响相互作用所形成的有机联系的反映。不同种的个体在空间联结程度的客观测定对研究两个种的相互作用和群落的组成及动态是有意义的, 对于认识生物群落中物种多样性的维持机制也有一定帮助, 同时, 种间联结测定还提供了一个客观认识自然种群的方法, 因而无论在理论上还是实践上都具有其重要意义。河南宝天曼自然保护区位于我国北亚热带向暖温带和第2级阶梯向第3级阶梯的过渡区, 特殊的地理位置为该区植物区系的形成提供了优越的历史地理条件, 决定了其南北交汇, 东西兼容的特点, 从而使该区成为植物群落类型比较复杂, 生物多样性相对丰富的地区之一。同时, 相对较少的人类活动干扰使该区保留了我国中部地区现存不多的天然次生森林资源。该区的研究基础比较薄弱, 仅限于综合考查和植物区系分析(史作民等, 1996; 宋朝枢, 1994), 其它方面的研究未见报道。本文对宝天曼保护区海拔1150~1500 m范围落叶阔叶林的20种主要乔木和26种主要灌木(含层间植物)种群的种间联结特征进行了初步研究。
1 研究地区自然条件和植被概况研究地区自然条件和植被概况见有关参考文献(史作民等, 1996; 宋朝枢, 1994)。
2 研究方法 2.1 样方设置本研究主要采用样方法取样, 乔木样方20 m×20 m, 每个乔木样方内设置5个2 m×2 m的灌木样方。调查记录乔木的种类、数量、高度、胸径、基径、冠幅, 灌木的种类、数量、高度、盖度, 同时调查记录各样地的海拔、坡度、坡向。共设置调查了28个乔木样方, 140个灌木样方, 计有乔木20种, 灌木(含层间植物) 26种(除去仅出现于1个样方内的种)。全部野外工作于1995年6~8月完成。
2.2 种间联结测度方法建立2×2联列表, 具体方法见相关参考文献(王伯荪等, 1985)。
2.2.1 χ2检验/χ2统计量度量由于取样为非连续性取样, 因此, 非连续性数据的χ2值用Yates的连续校正公式计算
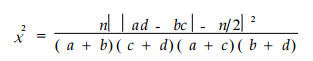
|
式中, n为取样总数。当χ2 < 3.841时, 种间联结独立; 当χ2≥6.635时, 种间有显著的生态联结; 当3.841≤χ2 < 6.635时, 种间有一定的生态联结。χ2本身没有负值, 因此判定正、负联结的方法是:若a> (a+b) (a+c) /n (即ad>bc)为正联结, 反之为负联结(王伯荪等, 1985; 郭志华等, 1997; Whittaker, 1986)。
2.2.2 共同出现百分率PC共同出现百分率PC也是用来测度物种间正联结程度的。其计算公式为:
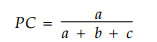
|
PC的值域为[0, 1]。其值越趋近于1, 则表明该种对的正联结越紧密(王伯荪等, 1985; 郭志华等, 1997; Whittaker, 1986)。
2.2.3 联结系数AC联结系数AC用来进一步检验由χ2所测出的结果及说明种间联结程度。其计算公式如下:

|
AC的值域为[-1, 1]。AC值越趋近于1, 表明物种间的正联结性越强, AC值越趋近于-1, 表明物种间的负联结性越强; AC值为0, 物种间完全独立(王伯荪等, 1985; 郭志华等, 1997; Hurlbert, 1969)。
3 结果与讨论 3.1 宝天曼落叶阔叶林乔木种群种间联结宝天曼落叶阔叶林乔木种群间χ2统计量数阵、共同出现百分率半矩阵图以及联结系数半矩阵图分别见表 1和图 1。

|
|
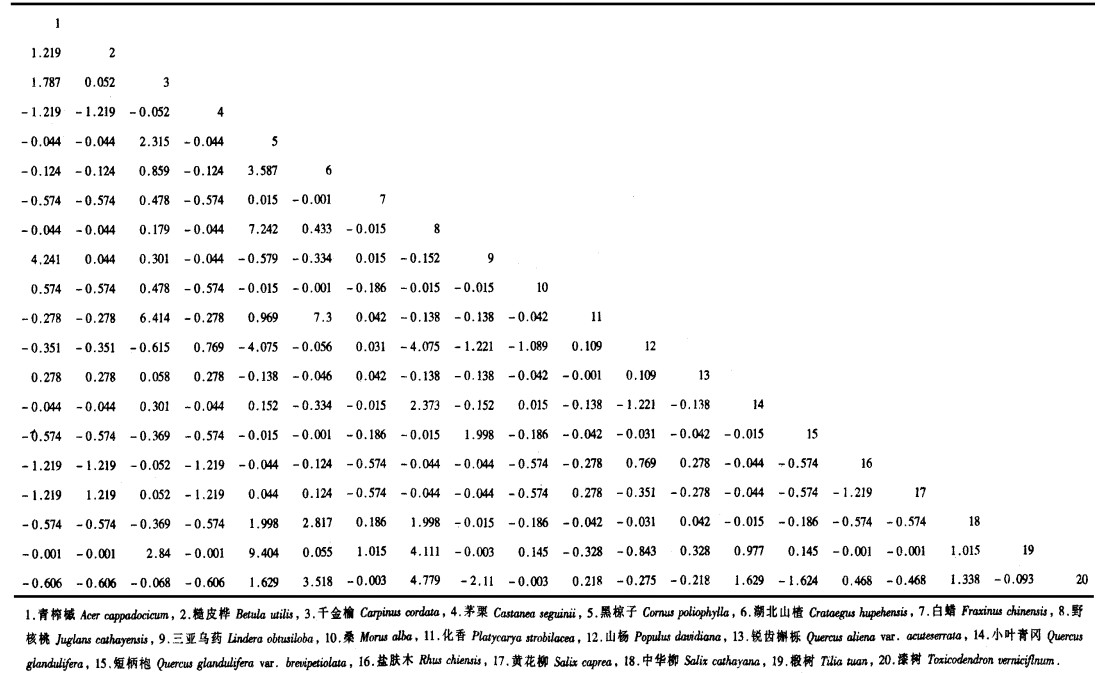
种间联结测定值本身具有种群生态学特征, 它在一定程度上衡量了种间的相互关系和植物对环境综合生态因子反应的差异。种间联结系数高表明一个种的存在对另一个种有利、或是这两个种对环境的差异有相似的反应; 相反, 种间联结系数低或负值则说明这两个种所需的环境条件不同或是一个种存在对另一个种不利而排斥它(王伯荪等, 1985)。
由表 1和图 1可见, 黑椋子和野核桃、黑椋子和椴树之间有明显的正联结, 是因为两者对环境有趋同反应或前者的存在对后者有利。黑椋子和山杨、野核桃和山杨之间有明显的负联结, 这主要由于它们对环境的要求不同所造成的。事实上, 它们的生境有明显的差异, 山杨主要分布在相对干旱的山坡, 为落叶阔叶林的先锋树种, 而黑椋子和野核桃主要分布在比较阴湿的沟谷, 土壤、阳光、水分等条件均有所不同。
3.2 宝天曼落叶阔叶林灌木种群种间联结宝天曼落叶阔叶林灌木种群间χ2统计量数阵、共同出现百分率半矩阵图以及联结系数半矩阵图分别见表 2和图 2。

|
|
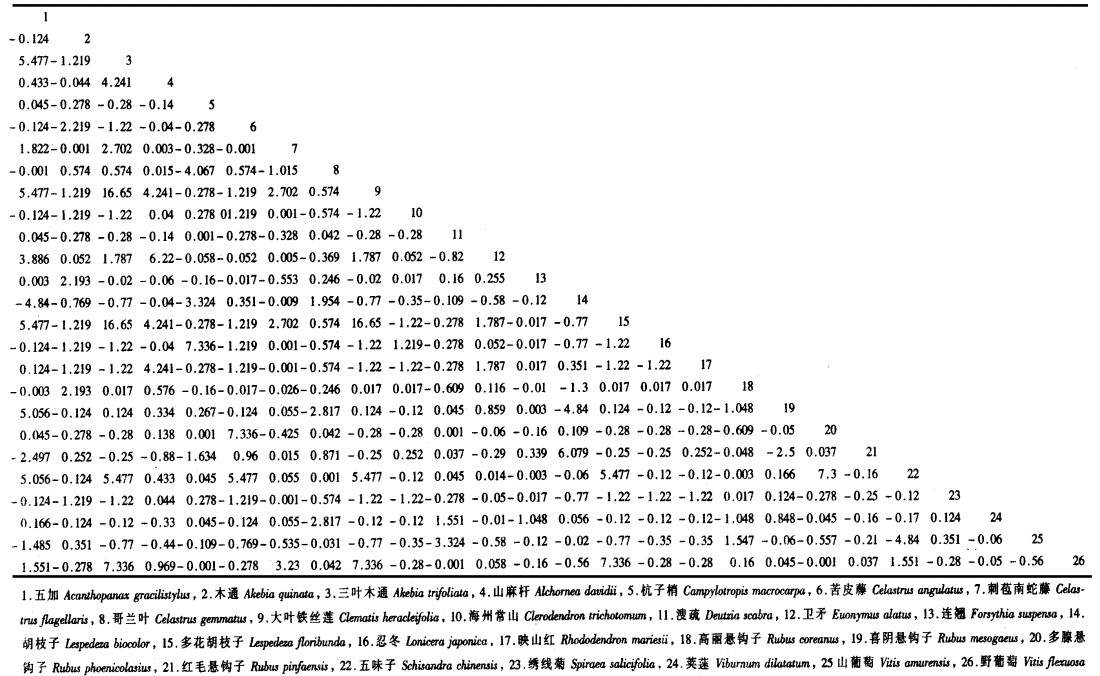
由表 2和图 2可见, 三叶木通、大叶铁线莲和多花胡枝子两两之间存在明显的正联结, 这主要归因于3者对环境有相似的要求。事实上, 这3个种具有相似的生物生态学特征, 喜欢阳光相对充足, 土壤不很阴湿的环境。五加和胡枝子、喜阴悬钩子和胡枝子、五味子和山葡萄之间存在较明显的负联结, 这主要由于它们对生境条件有不同的要求。五加、喜阴悬钩子和五味子喜欢相对阴湿的环境, 而胡枝子和山葡萄在相对干旱的环境生长良好, 具有较强的耐旱性。
3.3 种间联结与生态位重叠①物种间的联结性与其生态位重叠之间有较大的相关性, 种间联结高的种对, 一般有高的生态位重叠(彭少麟等; 1990)。宝天曼落叶阔叶林乔木种对黑椋子和椴树、黑椋子和野核桃、青榨槭和三亚乌药间均有显著或一定的正联结, 它们之间的生态位重叠也很大, 分别为0.660、0.461和0.733 (Pianka 公式, 下同); 黑椋子和山杨以及野核桃和山杨之间有最强的负联结, 它们之间的生态位重叠也很小, 分别为0和0.132。灌木种对三叶木通和多花胡枝子、杭子梢和忍冬以及苦皮藤和五味子之间有显著或一定的正联结, 它们之间的生态位重叠也很大, 分别为1.000、0.845和0.833;五加和胡枝子、喜阴悬钩子和胡枝子以及五味子和山葡萄之间有最强的负联结, 它们之间不存在生态位重叠, 其生态位重叠值均为0。由此可见, 种群间的正联结性越强, 其生态位重叠值就越大; 反之, 种间的负联结性越强, 其生态位重叠值就越小。
① 史作民:河南宝天曼保护区植物群落多样性研究(博士生毕业论文)。
3.4 生态种组群落中生态习性相似的种可以联合为一生态种组(Mueller-Dombois et al., 1986)。群落内的种间联结性揭示了群落中不同种类因受小生境因子影响而体现在空间分布上的相互关系(扬一川等, 1994)。以负联结性作为划分种组的界限, 同一种组内两两之间有尽可能大的正联结为原则, 本文对宝天曼落叶阔叶林主要乔木种群进行了生态种组划分, 大致可将20个乔木种群划分为4个生态种组。
第1生态种组为黑椋子、野核桃和小叶青冈, 该种组内两两之间的χ2值、AC值和PC值都较高, 正联结性很强。它们倾向生长在一起, 都喜欢较为阴湿的环境, 对水分条件要求较高。第2生态种组有千金榆、化香、锐齿栎、椴树和漆树等, 该种组内两两之间多存在一定的正联结, 而且种组内各物种除了与其它少数物种间存在一定的负联结外, 与其它多数物种间有一定的正联结。该种组内的物种为中生树种, 它们不仅数量多, 而且分布广、生态幅度大, 多为群落的建群种或主要伴生种。湖北山楂、白蜡和山杨为第3生态种组, 种组内两两之间存在一定的正联结, 种组内的各物种除了与其它少数物种存在明显的正联结外, 与其它多数物种存在明显的负联结, 或联结不明显。种组内各物种为阳生树种, 在种间竞争中处于劣势, 随着群落的进一步发育将逐渐被淘汰。余下的青榨槭、糙皮桦、茅栗、三亚乌药、桑、短柄、盐肤木、黄花柳和中华柳等为第4生态种组, 种组内除青榨槭和糙皮桦、青榨槭和三亚乌药、糙皮桦和三亚乌药、黄花柳和糙皮桦之间有明显的正联结外, 其它两两之间都表现为相互独立, 而且大多与其它各物种间都表现出相互独立。这些物种可以分为两种类型, 桑、盐肤木、黄花柳和中华柳等为植物群落的次要伴生种, 数量较少, 呈随机分布; 青榨槭、三亚乌药、糙皮桦、茅栗和短柄等的数量也较少, 但分布比较集中, 与其它物种的联结关系较弱。
3.5 种间联结性与植物多样性保护种间联结性是对一定时期内植物群落组成物种之间相互关系的静态描述, 因此对于顶极群落而言, 群落组成物种之间的联结性变化不大; 但对于非顶极群落, 由于随着群落的动态发育过程, 群落的组成物种在群落不同发育阶段会出现此消彼长的动态变化, 各物种的群落功能及其相互关系也会随之发生变化, 种间联结性将发生或多或少的变化。虽然种间联结性是对物种之间静态关系的描述, 但这种关系不仅包括空间分布关系, 同时也隐含着物种之间的功能关系。因此, 种间联结性研究对于认识特定群落内珍稀濒危物种的群落结构和功能地位以及其它物种与之相互关系具有一定作用, 同时在一定程度上, 该研究对于寻求特定群落内的关系物种也具有一定帮助。总之, 种间联结性研究对于保护植物群落稳定性, 从而保护植物群落多样性具有重要作用, 同时, 种间联结性研究对于特定物种的保护也有比较重要的作用, 亦即可以通过寻找和保护与之正联结性较强的物种来保护特定物种的生存环境, 最终达到保护特定物种的目的。
4 结论宝天曼落叶阔叶林黑椋子和野核桃、黑椋子和椴树乔木种群之间有明显的正联结, 黑椋子和山杨、野核桃和山杨乔木种群之间有明显的负联结; 三叶木通、大叶铁线莲和多花胡枝子灌木种群两两之间存在明显的正联结, 五加和胡枝子、喜阴悬钩子和胡枝子、五味子和山葡萄灌木种群之间存在较明显的负联结。
在宝天曼落叶阔叶林进行物种联结性研究时, 以χ2检验为基础, 结合联结系数AC和共同出现百分率PC等指标来分析可以得到较好的结果。
种间的正联结性越强, 其生态位重叠值越大; 种间的负联结性越强, 其生态位重叠值越小。
根据种间联结特征, 将宝天曼落叶阔叶林主要乔木种划分为4个生态种组。第1生态种组为黑椋子、野核桃和小叶青冈, 第2生态种组为千金榆、化香、锐齿栎、椴树和漆树, 湖北山楂、白蜡和山杨为第3生态种组, 第4生态种组为青榨槭、糙皮桦、茅栗、三亚乌药、桑、短柄、盐肤木、黄花柳和中华柳等。
郭志华, 等. 1997. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究. 植物生态学报, 21(5): 424-432. DOI:10.3321/j.issn:1005-264X.1997.05.004 |
彭少麟, 王伯荪.鼎湖山森林群落优势种群生态位重叠研究, 见: 热带亚热带森林生态系统研究(6集).北京: 科学出版社, 1990
|
史作民, 刘世荣, 王正用. 1996. 河南宝天曼种子植物区系特征. 西北植物学报, 16(3): 329-335. DOI:10.3321/j.issn:1000-4025.1996.03.020 |
宋朝枢主编.宝天曼自然保护区科学考察集.北京: 中国林业出版社, 1994
|
王伯荪, 李鸣光, 彭少麟著.植物种群学.广州: 中山大学出版社, 1989
|
王伯荪, 彭少麟. 1985. 南亚热带常绿阔叶林种间联结测定技术研究Ⅰ.种间联结测式的探讨与修订. 植物生态学与地植物学丛刊, 9(4): 274-285. |
杨一川, 等. 1994. 峨嵋山峨嵋栲、华木荷群落研究. 植物生态学报, 18(2): 105-120. DOI:10.3321/j.issn:1005-264X.1994.02.001 |
Greig P-Smith. 1983. Quantitative Plant Ecology. Oxford: Blackwell Scientific Publication.
|
Hurlbert S H. 1969. A coefficient of interspecific association. Ecology, 50: 1-9. DOI:10.2307/1934657 |
Mueller-Dombois D., Ellenberg H.著(鲍显诚等译).植被生态学的目的和方法.北京: 科学出版社, 1986
|
Whittaker R H著(王伯荪译).植物群落排序.北京: 科学出版社, 1986
|