 2001, Vol. 37
2001, Vol. 37文章信息
- 潘开文, 刘照光.
- Pan Kaiwen, Liu Zhaoguang.
- 连香树人工幼林群落营养元素含量、积累分配和循环
- THE DISTRIBUTION AND CIRCULATION OF NUTRIENT ELEMENT CONTENTS AND ACCUMULATION IN THE YOUNG ARTIFICIAL CERCIDIPHYLLUM JAPONICUM COMMUNITY
- 林业科学, 2001, 37(2): 1-12.
- Scientia Silvae Sinicae, 2001, 37(2): 1-12.
-
文章历史
- 收稿日期:1999-06-23
-
作者相关文章


连香树(Cercidiphyllum japonicum)为国家二级珍稀濒危保护植物(宋朝枢等, 1989), 属于连香树科连香树属, 是东亚植物区系的特有种, 产于日本和中国。连香树在我国残遗分布在暖温带及亚热带山地的杂木林中, 常与水青树(Tetracentron sinense)、珙桐(Davidia involucrata)、银雀树(Tapiscia sinensis)等古老树种伴生。由于连香树为白垩纪残遗树种, 在植物分类系统上有重要的科学价值, 所以, 前期有关连香树的研究报道主要集中于其系统演化问题的探讨(Swamy et al., 1949; Metcalfe et al., 1957; Erdtman, 1966; Davis, 1966; Ratter et al., 1973; 王东, 1990a, 1991b)。近年来, 人们发现连香树是速生优良的用材和观赏树种, 又是重要的中药材和香料工业植物(宋朝枢等, 1989), 进而逐渐被世界各国作为庭院绿化树种广泛引种栽培(Gibbons, 1995)。中国科学院成都生物研究所于“七五”期间在山地常绿、落叶阔叶混交林和针阔混交林区域营建大面积连香树林, 并研究报道了其生物量特征和与土壤活性铝的关系(潘开文等, 1999a, 1999b)。本文主要探讨连香树幼林群落营养元素的含量、分配和循环特征, 为进一步将连香树作为其分布区域的造林树种以恢复退化的森林提供理论依据和技术借鉴。
1 试验区概况试验区地处青藏高原东南边缘四川省境内的中国科学院茂县生态站, 地理位置103°54′04″~103°56′52″E和31°37′20″~31°44′53″N; 地貌为青藏高原东南边缘深切割, 海拔1554.9~4200.0 m, 气候复杂多样, 植被、土壤垂直带谱明显。成土母岩为志留系千枚岩、泥盆系灰岩、千枚岩夹薄层石英岩和寒武系变质岩等。
试验点在海拔2100~2300 m之间, 土壤为棕壤。年降水量900~1100 mm。自然次生植被主要为辽东栎(Quercus liaotungensis)、虎榛(Ostriopsix spp.)、小果蔷薇(Rosa cymosa)等落叶灌丛, 人工种植的林木主要有油松(Pinus tabulaeformis)、华山松(Pinus armandii)、落叶松(Laris spp.)、云杉(Picea spp.)和连香树等。
2 研究方法 2.1 生物量调查及植物样品的收集在环境条件基本一致的10年生(含苗期)连香树人工群落中, 随机每木检尺60株, 统计其平均胸径和树高。采取标准木法, 选取3株标准木进行树干解析。同时, 采用收获法, 地上部分按水平0.5 cm、垂直1.0 m为长度单位, 地下部分按水平0.40 m、垂直0.10 m为长度单位分层分器官收获; 地下部分用土壤筛和游标卡尺分径级称重, 然后分器官分层取少量样品烘干折算其干重。另分别选择2株最大和最小株, 并随机取不同径级的连香树8株, 采用收获法分器官分层测其生物量。
连香树生长的主要季节在6~9月, 本研究取植物活性样品供室内化学分析的时间在8月和9月, 分两次进行, 第1次, 与生物量调查同步进行, 分器官分层次分别从上述15株连香树中获取, 从样株中取的每一器官尽量均匀, 以能代表该样株该器官的特性, 如树干取样部位包括树基、胸径处和树梢等3处的混合, 大径级的树根和侧枝分别包括各径级根和枝的样品的混合等; 第2次, 在样地中随机取30处, 每处包括上述不同器官层次和根与枝级的样品。为了尽可能节省费用, 在室内, 将每次取回的样品按器官、径级和层次等分类, 每类等量均匀混合组建供室内化学分析的样品。
在上述连香树群落中, 随机布设10个1 m×1 m的木框架尼龙纱漏斗式收集器, 逐月收集枯落叶, 以烘干重累计(温肇穆等, 1991)。并按比例取逐月收集的样品供室内化学分析。在叶凋落完后, 清理地上残存的枯落物, 称重并分枯枝和落叶取混合样烘干称重以折算其干重和养分。
2.2 土壤调查及其样品的收集在上述连香树群落中, 随机布设4个土壤剖面, 调查其土壤剖面的特征, 取少量土壤(0~40 cm)供室内化学分析。
2.3 连香树群落内外降雨测定在上述样地中, 按每径级选1~2株树, 用聚乙烯管一端固定在树干上(约离树干基部1 m), 然后在树干上螺旋式围绕3~4圈, 用沥青粘牢, 最下面一端放入靠近树干基部的接收筒中测定茎流(冯宗炜等, 1985)。在林内随机布设10个标准雨量筒收集穿透林冠水, 林外随机布设3个标准雨量筒收集林外降水。每次下雨后即量测, 并取样迅速带入室内, 放入冰箱中冷藏。无论下雨否, 只要有样品, 每隔1个月即进行水化学测试, 在测试时, 将该月收集的雨水按每次收集量的多少, 按比例取样混合进行化学分析。观测时间2 a。
2.4 样品化学测定 2.4.1 土壤有效N用扩散吸收法, 全N用半微量开氏法; 有效P用碳酸氢钠浸提钼锑抗比色法, 全P用钼锑抗比色法; 有效K用乙酸铵浸提火焰光度法, 全K用氢氟酸-高氯酸消煮-火焰光度法; 有效和全量Ca、Mg、Fe、Mn都用原子吸收光谱法, 活性和全Al用氟化钾取代EDTA容量法(刘光崧, 1996)。
2.4.2 植株全量N用重铬酸钾硫酸消化法, P用钒钼黄比色法, K用火焰光度法, Ca、Mg、Fe、Mn都用原子吸收光谱法, Al用氟化钾取代EDTA容量法(刘光崧, 1996)
2.4.3 水样全N用半微量开氏法, 全P用钼锑抗比色法, 全K用火焰光度法, Ca、Mg、Fe、Mn都用原子吸收光谱法, 全Al用氟化钾取代EDTA容量法(张宏陶, 1989)。
3 结果与分析 3.1 植物器官营养元素含量分布连香树群落各营养器官中N元素的平均含量分别为叶(13.17 g·kg-1, 单位下同) > 皮(6.70) >根(6.22) >枝(4.60) >去皮干(2.00) (表 1), 其极差为11.17, 叶中N含量是去皮干的5.6倍。侧枝和根的径级越小, 其N含量越小, 反之则相反; 如在 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm的根中, 其N含量分别为3.10、3.20、3.90、8.40;在≥1 cm和 < 1 cm的侧枝中的N含量分别为3.80和5.40 (表 1)。植物器官的取样部位的不同也影响器官中的N含量, 如在 < 4 m、4~6 m和 > 6 m高度的叶中N含量分别为13.00、13.10和13.17 (表 2), 这种结果和N进入植物体结合成氨基酸和合成蛋白质的生理功能有关(拉夏埃尔, 1985)。
|
|

|
|

P元素的平均含量规律为叶(0.97) > 皮(0.78) >侧枝(0.57)、根(0.57) >去皮干(0.33), 其极差为0.64, 叶中P的含量是去皮干的1.9倍。在 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm级的根中, P含量分别为0.43、0.44、0.51和0.89;在 < 4 m、4~6 m和 > 6 m高度范围内的叶中, P含量分别为0.98、0.91和1.03;反映了根级越大, 叶的光合代谢越弱, 含P量越少, 反之则相反, 只不过其变幅不及N元素更明显(表 1、2)。
K元素的平均含量规律为叶(7.64)皮(3.24) >根(3.68) >侧枝(2.70) >去皮干(0.84), 叶中K元素的含量是去皮干的9.1倍, 其极差为6.80。在不同径级侧枝中, > 1 cm的侧枝含K量(1.49)仅是小于1 cm侧枝(3.90)的0.38倍; 在 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm的径级根系中, K含量分别为2.57、2.99、3.15、7.30;在 < 4 m、4~6 m和 > 6 m高度的叶中K含量分别为7.14、7.22和8.55, 表明营养器官代谢越强的部位, K含量越高(表 1、2)。
Ca元素的平均含量规律为叶(28.71) > 皮(24.43) >根(15.55) >侧枝(9.84) >去皮干(6.71), 叶中Ca含量是去皮干的4.3倍, 其极差为22.00。在侧枝中, > 1 cm和 < 1 cm的侧枝含Ca量分别为7.74和11.93;在 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm径级的根系中, Ca含量分别为7.76、7.84、8.27和8.4;在 < 4 m、4~6 m和 > 6 m高度范围内的叶中Ca含量分别为26.40、29.00和30.72。表明Ca含量在代谢旺盛的部位高(表 1和2)。
Mg元素的含量分布规律为叶(2.55) >根(1.34) > 皮(0.78) >侧枝(0.57) >去皮干(0.00), Mg是叶绿素的构成元素, 因而叶中含Mg量高。在 > 1 cm和 < 1 cm的侧枝中, Mg含量分别为0.50和0.63;在 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm级的根系中, Mg含量分别为0.51、0.71、1.11和3.05;在 < 4 m、4~6 m和 > 6 m高度范围内的叶中, Mg含量分别为2.52、2.53和2.60。可见, Mg在代谢活动越旺盛的部位, 其含量越高(表 1、2)。
Fe元素的含量分布规律为根(2.48) >叶(0.23) > 皮(0.22) >侧枝和去皮干(0.12)。根中Fe平均含量是枝干的20.7倍, 其极差为2.36。Fe元素含量在植物体内的分布不同于N、P、K、Ca和Mg。代谢活动越弱的叶, Fe含量越高, 反之则相反, 如在 < 4 m、4~6 m和 > 6 m高度范围内连香树叶的Fe含量分别为0.32、0.20和0.17 (表 2)。在侧枝和根中, 代谢活动越强, Fe含量越高, 反之则相反, 这尤其在不同径级根系中表现得最为明显; 如 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm的根中Fe含量分别为0.26、1.04、1.90和6.72, 其极差为6.46, < 0.1 cm的根中Fe含量是大于1 cm根的25.8倍。Fe的这种含量分布规律和其在植物体中的功能相吻合。如根系中, 由于越细的根系吸收活动越频繁, Fe运载体配合物越多, 故其平均含Fe量越高; 而在叶中, 越下面的叶, 光合作用越弱, 呼吸作用越强, 磁性氧化铁和磷酸铁等含量越高, 故植物叶片代谢活动越强, 其含量越高, 反之则相反(王夔, 1992)。
Mn元素平均含量分布规律为根(0.26) >叶(0.03) > 皮、枝(0.01) >干(0.00), 其极差为0.26。叶中Mn含量与叶的部位关系不明显(表 2); 侧枝和根中的Mn则有在越细的器官中含量越高的态势, 如 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm的根中其Mn含量分别为0.05、0.20、0.50和0.30, 总体反映出代谢功能强的部位, Mn的含量高。
Al元素平均含量分布规律为根(13.85) >叶(1.21) > 皮(1.17) >干(1.04) >枝(0.73), 其极差为13.12, 根中Al的含量是枝中的18.97倍。在叶中, 光合作用越弱的部位, 含Al量越低; 在侧枝中, 越粗的侧枝含Al量越高; 而在根系中, 则器官越细, Al含量越高, 如 > 1 cm、0.5~1.0 cm、0.1~0.5 cm和 < 0.1 cm的根系中其Al元素含量分别是1.26、5.56、9.43和39.16。
各元素在植物各器官中的平均含量规律为Ca (15.55) >N (6.22) >Al (3.60) >K (3.68) >Mg (1.05) >P (0.64) >Fe (0.63) >Mn (0.06) (表 2), 明显有别于油茶群落中油茶各器官元素平均含量呈现出N>K>Mg>Ca>P的趋势的结论(谭云峰等, 1989), 这说明连香树对Ca、Al的需求更高于对P和K的需求。大多数植物含Al量仅0.00~0.5 g/kg, 而连香树植物群落平均含Al达3.60 g/kg, 反映了其富Al, 这和其原始生境亚热带中山酸性棕壤常绿阔叶与落叶混交林类型的优势植物富Al的结论一致(侯学煜, 1982), 但不富Mn。由于其含Al量高, 因而是酸性指标植物(侯学煜, 1982)。8种元素在各器官中的总含量表现为叶(54.51) > 皮(37.33) >根(35.23) >枝(19.07) >干(11.04), 而在根系中又表现出(>1.0 cm)侧根(15.94) < (0.05~1.0 cm)侧根(21.97) < (0.1~0.5 cm)侧根(28.77) < (< 0.1 cm)侧根(74.23)。表明器官代谢越旺盛, 各元素的浓度越大, 反之则相反; 但是, 各器官中的不同元素因其具有不同的生理功能, 故其含量规律又有特殊性, 如干中Al含量高、代谢活动弱的叶中Fe含量高等(表 1、2)。
3.2 连香树群落营养元素的分配N元素在各营养器官的分配规律为:根(164.45 kg/hm2, 单位下同) >叶(126.76) >去皮干(83.89) >枝(70.63) > 皮(61.26);P:根(22.01) >去皮干(13.84) >叶(9.34) >枝(8.75) > 皮(7.13);K的为根(317.79) >叶(73.54) >枝(41.46) >去皮干(35.23) > 皮(29.63);Ca:根(374.67) >去皮干(281.45) >叶(276.33) > 皮(223.39) >枝(151.08);Mg:根(33.35) >叶(24.54) >枝(8.75) > 皮(7.13) >去皮干(0.00);Fe:根(36.45) >去皮干(5.03) >叶(2.21) > 皮(2.01) >枝(1.84);Mn:根(5.05) >叶(0.29) >枝(0.15) > 皮(0.09) >去皮干(0.00);Al:根(195.15) >去皮干(43.62) >叶(11.65) >枝(11.21) > 皮(10.70) (表 3)。可见, P和Al、Ca和Fe在各营养器官中的分配趋势一致, 即分别表现为根>去皮干>叶>枝 > 皮和根>去皮干>叶 > 皮>枝; Mg和Mn的分配趋势一致, 即根>叶>枝 > 皮>去皮干; N和K在各营养器官的分配趋势相近, 即根>叶>枝≠去皮干≠皮, 表明上述各组元素间彼此有着更紧密的相关性。
就8种元素在各营养器官中的总量而言, 根(968.92) >叶(524.66) >去皮干(463.06) > 皮(341.34) >枝(293.87), 根系中的元素占总量的37.38%。就植物体中分配的各元素而言, 则表现出Ca (1306.92) >N (506.99) >K (317.65) >Al (272.33) >Mg (73.77) >P (61.07) >Fe (47.54) >Mn (5.58), Ca是N的2.58倍、K的4.11倍、P的21.40倍, 表明该植物是富Ca植物; Al是P和N、K的4.4、0.54和0.86倍, 也表明其是富Al植物, 只不过不及富Ca强烈。植物体中Ca和Al等有拮抗作用(孙羲, 1980), 连香树对Ca的高度富积可能与其富Al而消去过多的Al离子的毒害有关(表 3)。
|
|

由于连香树为落叶树种, 每年长出的树叶在当年12月前全部凋落, 所以, 按国内外普遍采用的算法, 连香树的存留是指该树种中除叶以外的器官中的元素总量之和, 即各器官生物量乘以相应元素的浓度, 结果见表 4。连香树幼林对8种元素的积累量为2067.19 kg/hm2, 年均存留量为206.72 kg/ (hm2·a)。连香树群落对Ca的存留最多, 为103.06 kg/ (hm2·a) (单位下同), 占8种元素存留量的49.85%, 其次分别为N (38.03) >Al (26.07) >K (24.41) >P (5.17) >Mg (4.92) >Fe (4.53) >Mn (0.53)。因此, 连香树群落对Ca的富集作用最强, 对P和Mn的富集作用较小, 而对Al的富集作用相对较强, 大于植物生长的一些大量元素。这种富积特性和其所处的原始植被类型中优势植物的富积特性一致(侯学煜, 1982), 表明富积作用是植物、气候和土壤三相的统一。
|
|
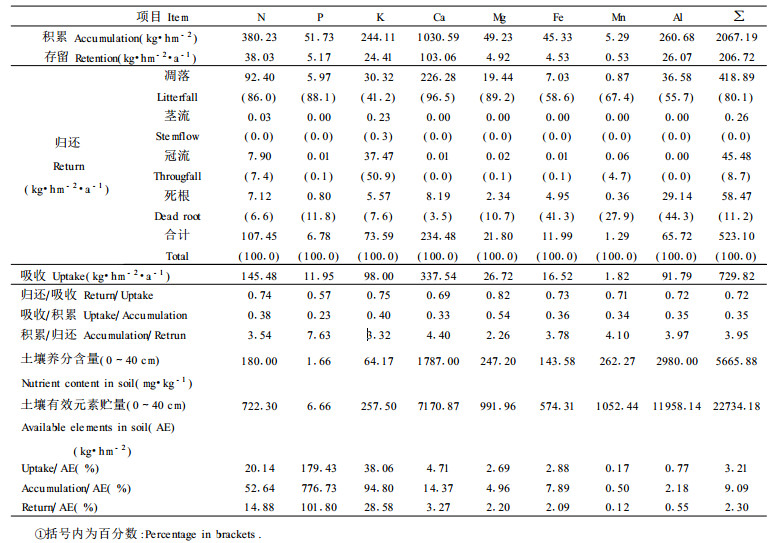
归还含3种:凋落物归还, 降水淋溶归还, 死根归还。
(1)凋落物归还 由于连香树为10年生, 且为落叶树种, 所以, 其归还的凋落物主要为树叶。凋落物量乘以相应元素的含量即为凋落物归还的元素量(表 4)。8种元素通过凋落叶归还量为418.89 t/ (hm2·a), 占年归还量的80.1%, 表明凋落叶是养分元素归还于土壤的主要归还途径; 然而对K元素而言, 通过凋落叶归还的量仅占K归还总量的41.2%, 表明凋落物对K而言不是最主要的归还途径(表 4)。
(2)降水淋溶归还 根据以下三式计算降水淋溶归还量(表 4)。

|
(1) |

|
(2) |

|
(3) |
式中, LAS和LAT分别为茎流(Leaching amount through stem)和冠流淋溶量(Leaching amount through throughfall); CNES和CNEP分别为茎流养分浓度(Concentration of nutrient elements in stemflow)和空旷雨养分浓度(Concentration of nutrient elements in precipitation); CNET为冠流养分浓度(Concentration of nutrient elements in througfall); LA、S和T分别表述淋溶量(Leaching amount)、茎流(Stemflow)和冠流(Throughfall)。
降水茎流和冠流归还量分别为0.26和45.48 t/ (hm2·a), 分别占降水淋溶归还量的0.57%和99.43% (表 4), 表明在降水归还途径中, 主要是冠流归还, 而茎流归还量小得几乎可以忽略不计。降水淋溶归还量为45.48 t/ (hm2·a), 占总归还量的8.7%, 与叶占年归还量的80.1%相比(表 4), 降水淋溶归还是极次要的途径。Al几乎不通过淋溶归还, P、Ca、Mg、Fe等淋溶量少, 但淋溶归还的K、N分别占其对应元素总归还量的50.9%和7.4%;表明淋溶归还的N占一定比例, 而对K而言, 淋溶归还是主要的归还途径, 这与K以离子存在于植物体有关。降水淋溶损失可能主要来自细胞壁的蒸腾液, 而细胞原生质只是选择性地从液流中吸取了当时所需的营养元素, 其余的营养物质聚结在细胞壁和角质层内, 当降水时它们被氢离子(H+)所交换出来而随降水归还于土壤(冯宗炜等, 1985)。
(3)死根归还 假定在 < 2 mm级的细根中, 每年归还的细根占总细根生物量的25% (哈默特等, 1989), 根据其归还的生物量乘以对应元素的浓度得其通过死根归还于土壤的养分为58.47 kg/ (hm2·a), 占总归还量的11.2%, 因而是极次要的归还途径。但对Fe、Al而言, 其归还量分别占对应元素总归还量的41.3%和44.3%, 故对Fe、Al而言仍是重要的归还途径(表 4)。
3.3.3 循环特征表 5显示, 10年生连香树群落积累了N、P、K、Ca、Mg、Fe、Mn、Al等8种元素260.68 kg/hm2, 且Ca (1030.59 kg/hm2, 单位下同) >N (380.23) >Al (260.68) >K (244.11) >P (51.73) >Mg (49.23) >Fe (45.33) >Mn (5.29);年存留总量为206.72 kg/ (hm2·a), 其中, Ca[103.06 kg/ (hm2·a), 单位下同]>N (38.03) >Al (26.07) >K (24.41) >P (5.17) >Mg (4.92) >Fe (4.53) >Mn (0.53)。年归还量为523.10 kg/ (hm2·a), 是存留量的2.53倍。各元素归还量表现为Ca[234.48 kg/ (hm2·a), 单位下同]>N (107.45) >K (73.59) >Al (65.72) >Mg (21.80) >Fe (11.99) >P (6.78) >Mn (1.29), 与元素存留量的大小顺序基本一致。年吸收量为年存留量与年归还量之和(Twilley et al., 1986), 连香树群落年吸收量为729.82 kg/ (hm2·a), 其中有吸收量的28.32%存留在立木上, 57.40%通过凋落、6.27%通过降水淋溶、8.01%通过死根等途径归还, 归还量为吸收量的71.68%。各元素年吸收量表现为Ca (337.54) >N (145.48) >K (98.00) >Al (91.79) >Mg (26.72) >Fe (16.52) >P (11.95) >Mn (1.82), 其吸收量大小顺序和归还量大小顺序完全一致, 与存留量的基本一致(表 4)。植物对其结构组成物质Ca的吸收量最大, 分别是N的2.3倍、K的3.4倍、Al的3.7倍、Mg的12.6倍、Fe的20.4倍、P的28.2倍、Mn的185.5倍。对植物生长的大量元素N、P、K而言, 群落对P的吸收极少, 仅分别为N和K的0.08及0.12倍, 且远低于对Al、Mg、Fe的吸收量; 吸收最少的是Mn。植物本身的选择吸收与气候、土壤和物种有关, 连香树群落对P吸收较少的原因可能与土壤中有效P较缺乏有关(表 4)。土壤中其余有效元素都相对较充足, 故连香树群落对上述各元素的吸收与富集特性是其本身的物种选择吸收特性。连香树群落中各元素有自己的循环特征, 可通过周转期、利用系数和循环系数表示(林鹏等, 1985)。
|
|

元素归还量与吸收量的比值称为归还比, 表征元素的循环强度(Duvigneaud, 1982), 有人亦称为循环系数(林鹏等, 1985)。某元素的归还比越大, 循环强度越大, 林木生长对土壤库存元素的耗费越小, 越有利于该元素在土壤中的积累(温肇穆等, 1991)。10年生连香树群落平均归还比为0.72;其中Mg的归还比最大, 为0.82;最小为P元素, 归还比为0.57。各元素归还比大小顺序为Mg (0.82) >K (0.75) >N (0.74) >Fe (0.73) >Al (0.72) >Mn (0.71) >Ca (0.69) >P (0.57) (表 4)。表明, 最有利于土壤中Mg的积累; 其次是K、N、Fe、Al、Mn; 最不利于在土壤中积累的元素是P。
元素吸收量与积累量的比值, 称为利用系数(Utilization coefficient) (陈灵芝, 1983)。林木对元素的吸收量是维持其生长所需要的元素量, 但是, 吸收进入植物体中的元素并非全部被利用而积累, 因此元素利用系数越小, 林木对该元素的利用效率越高, 反之则相反。连香树群落对8种元素的利用效率都较高, 平均利用系数为0.35。利用效率最高的是P, 其利用系数为0.23;其次是Ca、Mn、Al、Fe、N、K, 利用系数为0.33~0.40;利用效率最低的是Mg, 其利用系数为0.54 (表 4)。元素的积累量与归还量的比值为元素的周转期(Circulation period) (Frank et al., 1982)。元素周转期与元素利用效率相辅相存, 周转期越长, 元素的利用效率越高, 反之则相反。8种元素的平均周转期为4 a左右。P为周转期最长, 为7 a左右, 其利用效率最高; 其次是Ca, 为4.5 a左右; 再次是Mn、Al、Fe、N、K, 为3~4 a左右; Mg的周转期最短, 不足3 a, 其利用率相对最低; 周转期分析结果与循环强度的分析结果一致。
土壤中的有效元素贮量表现为Al (11958.14 kg/hm2, 单位下同) >Ca (7170.87) >Mn (1052.44) >Mg (991.96) >Fe (574.31) >N (722.30) >K (257.50) >P (6.66) (表 4), 表明土壤中N、P、K有效贮存量较低, 这是由于酸性土壤中Al、Mn有效浓度极高, 不仅将N、P元素淋溶掉, 而且P与Fe、Al结合形成不溶性的磷酸铁(FePO4)和磷酸铝(AlPO4)。此外, 酸性土壤过酸对硝化细菌、共生和非共生的固氮细菌生长的不适宜, 亦使土壤中含K、P量较低。P和K含量低, Ca、Mg、Fe、Al等元素有效含量较高, 这与成土母岩为志留系千枚岩夹灰岩、泥盆系灰岩、千枚岩夹薄层石英岩、寒武系变质岩等成土母岩和上述母岩风化的残坡积物以及第四纪更新世次生黄土的成土母质有关; 还与pH值和氧化还原电位有关。显然这种含高Ca、Mg等的酸性土不同于含低Ca、Mg的亚热带酸性土(侯学煜, 1982)。
循环强度、利用系数和周转期都是从植物体中的元素的角度来表征元素的生物循环特征, 缺乏与土壤有效元素的联系, 也难以说明对土壤中有效元素的利用状况。为此提出群落吸收系数、群落归还系数和群落积累系数。群落吸收系数指群落年吸收某元素量与土壤对应有效元素库存量的百分数, 因而它实际上反映了群落利用土壤中有效元素的效率以及土壤库存有效元素的相对数量多少。吸收系数越大, 说明群落利用土壤库存有效元素的效率高, 土壤库存有效元素相对数量越小, 反之则相反。吸收系数一般小于100%, 若>100%, 则表明土壤中的某有效元素已不能满足植物生长的需要而成为限制因子。连香树群落吸收系数可分为四个等级:Mn、Al的吸收系数很低, 为0.17%~0.77%, 说明连香树群落对土壤库中的有效Mn、Al的利用率很低, 最低的是Mn; Ca、Mg、Fe的吸收系数为2.69%~4.71%, 表明连香树群落对土壤库中的Ca、Mg和Fe的利用率较低; 吸收利用率较高的是土壤中的K、N, 吸收系数为20%~39%;吸收利用率最高的是P, 吸收系数为776.73% (表 4), 表明土壤库中现有有效库存P尚不能满足连香树群落的吸收量, 而成为影响该群落发育的障碍因子, 因而要使其高产, 必须人工施用P肥。值得注意的是, 连香树群落对P的吸收量为11.95 kg/ (hm2·a), 而土壤库中能供其直接吸收利用的P仅是6.6 kg/ (hm2); 表明该群落对于改良土壤, 调节酸碱平衡, 促进矿物分解和P的活化有积极的贡献。群落归还系数指群落年归还量与土壤对应有效元素库存量的百分数, 因而其反映了群落归还量对土壤有效元素库存量的影响大小。群落归还系数越大, 则表明群落对土壤库中某有效元素的库存影响越大, 反之则相反。表 4表明, P的归还系数最大, 因而群落归还对土壤库中P的有效库存量影响最大; 其次, 群落归还对土壤库中K、N的有效库存量影响较大, 归还系数为14%~29%;群落归还对土壤库存其余有效元素量的影响大小表现为:Ca (3.27%) >Mg (2.20%) >Fe (2.09%) >Al (0.55%) >Mn (0.12%), 因而群落归还对土壤库存有效Ca、Mg、Fe、Al、Mn元素影响较小, 影响最小的是Mn。群落积累系数指群落积累量与土壤对应有效元素库存量的百分数, 其反映了群落利用土壤中有效元素的效率; 积累系数越大, 则表明群落利用土壤中有效元素的效率越高, 反之则相反。其和利用系数不同, 利用系数指群落对吸收后的元素的利用效率, 而不是指直接利用土壤库存有效元素的效率。表 4显示, 群落直接积累利用土壤库中有效元素的效率为P (776.73%) >K (94.80%) >N (52.64%) >Ca (14.37%) >Fe (7.89%) >Mg (4.96%) >Al (2.18%) >Mn (0.50%)。可见, 群落对土壤库中有效P的积累利用效率最高, 其次为K、N; 再次为Ca、Fe、Mg、Al; 对土壤中有效Mn的积累利用最低。显然, 群落积累系数与利用系数的分析结论不一致, 但与群落吸收系数的分析结论一致。
4 讨论与结论 4.1 降水淋溶归还的算法有的学者通过穿透林冠水和树干茎流水带下的元素量与空旷地降水元素量的差额来估算通过降水淋溶归还给土壤的元素量(Tsutsumi, 1982; 冯宗炜等, 1985; 温肇穆等, 1991), 据此计算出降水淋溶归还量如表 5所示。由于降水淋溶归还对连香树而言主要发生于其生长期(5~11月), 故以生长期降水量计算通过降雨淋溶归还量。表 5显示P、K、Ca、Mg、Mn通过降水淋溶而损失; 但N则通过降水, 使大气中的含N物质被吸附于叶表面而逐渐被叶吸收存留下来, 因而使N淋溶损失表现为负值, 这有悖于实际淋溶损失。该种方法估算的结果不符合连香树林的客观实际的原因在于忽略了叶吸收林冠截留的降水中所包含的营养物质。为避免上述方法的误差, 列出了本文中的计算方法。计算结果符合现实情况, 表明该计算方法既可行又合理。
4.2 几个系数的比较利用系数指元素吸收量与积累量的比值, 循环系数指元素归还量与吸收量的比值, 周转期指元素的积累量与归还量的比值, 3个系数都是描述群落对吸收后的元素的利用情况的指标。群落吸收系数指群落年吸收某元素量与土壤对应年有效元素库存量的百分数, 群落归还系数指群落年归还量与土壤年对应有效元素库存量的百分数, 群落积累系数指群落积累量与土壤年对应有效元素库存量的百分数, 因而它实际上反映了群落利用土壤中有效元素的效率和土壤库存有效元素的相对数量多少以及对土壤中某有效元素的库存的影响。可见, 利用系数、循环系数、周转期和群落归还系数、积累系数、吸收系数的显著区别在于:前三者描述群落吸收后的元素的循环特征, 着重强调植物对已吸收的元素的利用情况; 后三者描述元素在植物与土壤之间的循环特征, 重点描述群落吸收、利用土壤中有效元素以及对土壤中对应元素的影响的特征。
通常, 在利用系数、循环系数和周转期三个指标中, 周转期与循环系数的分析结果是一致的, 而与利用系数的分析结果无共同的可比性; 同样, 在群落归还系数、积累系数和吸收系数中, 群落积累系数与吸收系数的分析结果一致, 而与群落归还系数的分析结果无共同的可比性(表 4)。
4.3 落叶树种存留的算法 4.3.1 叶的元素迁移虽然连香树叶在不同生长季节、不同部位和不同器官中时刻都存在元素的体内迁移, 但本文由于仅为了计算吸收、归还和存留, 因而只研究叶凋落前后元素的迁移变化。凋落前用生长期叶元素平均含量(C1)、凋落后用凋落物叶中元素平均含量(C2)参与下式计算, 结果见表 6。

|
|
|
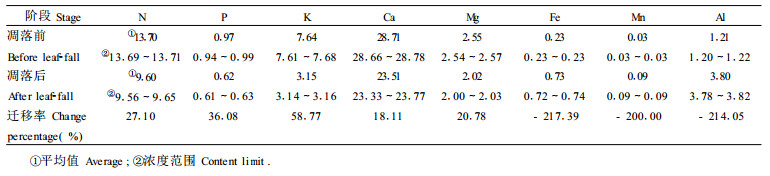
不同的元素迁移率不同(Althur et al., 1992)。8种元素的迁移率表现为Fe (217.39%) >Al (214.05%) >Mn (200.00%) >K (58.77%) >P (36.08%) >N (27.11%) >Mg (19.61%) >Ca (18.11%)。通常Fe在植物体中难转移, 而凋落前Fe的高度转移与其促进生理成熟有关, Al、Mn对植物的毒寒机理相近, 其高度转移主要是加快叶的死亡和脱落。K、P、N都是植物体中移动性较大的元素, 然而, K与P、N不同, 在植物体中, K主要以离子形态或可溶性盐类吸附在原生质的表面上, 而不是象P、N以有机化合物的形态存在, 因而其迁移率高于P、N。钙主要是细胞壁的结构成份, 它与中胶层果胶质形成钙盐而被固定下来, 不易转移和再利用, 因而其迁移率最低。不同的元素迁移方向也不同, 叶中的N、P、K、Ca、Mg等营养元素向其它器官迁移, 以减少其损失, 而Fe、Mn、Al则由其它器官向叶迁移, 以促成叶生理成熟而凋落归还林地。向体内迁移的元素往往是积累较多、需求量大的元素, 而向体外迁移的元素多数属积累和需求量较少的元素或是土壤库存有效量大的元素(表 3、6), 这种以损失需求量少或土壤库存量多的元素来换取需求量大或者土壤供应紧缺的元素的累积和再利用也是植物长期进化而适应环境的生理生存对策(Killingbeck et al., 1981)。凋落物中氮素含量的高低不仅影响到凋落物的矿化速度, 而且也反映出归还土壤的肥力效应和森林生态系统食物链中微生物的营养状况(卢俊培, 1994)。在连香树凋落物营养元素组成中, Ca (23.5 g/kg, 单位下同) >N (9.60>Al (3.80) >K (3.15) >Mg (2.02) >Fe (0.73) >P (0.62) >Mn (0.09) (表 6), 表明凋落物中N素含量高, 有利于微生物活动和矿化分解, 促进系统生物良性循环。
4.3.2 存留由于连香树为落叶树种, 每年长出的树叶在当年12月前全部凋落, 毫无疑问, 连香树的存留是指该树种中除叶以外的器官中的元素总量之和, 其算法是各器官生物量乘以相应元素的浓度。但是, 表 6已显示出, 连香树树叶在凋落前其养分已发生明显的转移, 虽然, 我们不能确切知道各元素转移到哪个具体部位, 但却知道各元素在体内和体外之间的转移, 向体外转移的即属于归还部分, 向体内转移的即属于积累和存留部分。所以, 在逻辑推理上, 存留应是在上述算法的基础上再加或减去因元素在叶与其它器官内外迁移而损失或增加的对应元素量。按此思路, 算出的元素循环特征如表 7所示。
|
|

将表 7和表 4的数据进行对照, 发现两种算法所得到的基本趋势较为一致, 如两表中的群落吸收系数、积累系数和归还系数从大到小排列顺序分别都为P>K>N>Ca>Fe>Mg>Al>Mn、P>K>N>Ca>Fe>Mg>Al>Mn和P>K>N>Ca>Mg>Fe>Mn>Al, 排列顺序完全一致。但两种算法所算出的系数排列顺序也有不尽一致的地方, 如表 7中各元素存留量排列顺序为Ca>N>K>Al>P>Mg>Fe>Mn, 而表 4中各元素存留量排列顺序为Ca>N>Al>K>P>Fe>Mg>Mn。两表中个别具体数据也有差异, 如表 4与表 7中8种元素积累的总量分别为2067.19 kg/hm2和2173.07 kg/hm2, 相对误差为5%。
总体而言, 两种算法的结果多数较为接近。前种算法为国内外普遍采用, 但本文的研究也发现了落叶树种叶养分的转移对此算法的影响, 如果落叶树种养分转移率更大, 那么误差可能就更大。所以, 作者倾向于后者的算法。
4.4 连香树群落的一些生理生态特性在8种元素中, 除Fe和Al外的其余元素在连香树各营养器官中的含量规律都表现为:代谢越旺盛的部位, 其含量越高。P和Al、Ca和Fe在各营养器官中的积累趋势一致, Mg和Mn的积累趋势一致, N和K的积累趋势相近, 表明上述各组元素间彼此有着更紧密的相关性(表 3)。8种元素中, 连香树对Ca的积累量最多, 这一方面是土壤中活性Ca含量高, 另一方面也是连香树对Ca的需求多, 因Ca和Al有拮抗作用, 这正好能消去过多的Al离子的毒害(表 3); 连香树对Al的大量富集与土壤中含大量活性Al有关, 因大量活性Al对植物有毒害作用, 因此, 连香树对Al的大量富集是否是其固有的生态生理特性尚值得进一步研究, 但连香树因Ca和Al的相克作用, 使连香树具有耐Al性。这个结论从理论上验证了连香树分布于亚热带富铝的酸性土上的合理性和可行性。
连香树群落中8种元素的主要归还途径因不同的元素而有所不同, K的主要归还途径是降水淋溶, 占归还总量的51.2%;其余元素主要的归还途径是凋落物, 占总归还量的55.7%~96.5%;Al几乎不通过淋溶归还, P、Ca、Mg、Fe等淋溶量少, N淋溶归还占总的7.4%;Fe、Al通过死根的归还量分别占总的41.3%和44.3% (表 4)。这些结论与元素的特性以及其在植物中的积累分布和移动的特点相吻合。
在8种元素中, 连香树群落利用效率最高的是P, 其次是Ca、Fe、Al、Mn、N、K, 最低的是Mg; 周转期最长的是P, 其次是Ca、Al、N、Fe、Mn、K, 最短的是Mg。显然, 周转期分析结果与循环强度的分析结果一致(表 4)。连香树群落吸收系数最小的是Mn、Al, 较小的是Ca、Mg、Fe, 较大的是K、N, 最大的是P。归还系数最大的是P, 较大的是K、N。积累系数最大的是P, 其次为K、N; 再次为Ca、Mg、Fe、Al, 最小的是Mn。显然, 群落积累系数与利用系数的分析结论不一致, 但与群落吸收系数的分析结论一致。
上述结果之间可互相佐证和解释, 比如土壤中P较缺乏, 群落积累系数与归还系数都大; 植物吸收P后, 对P的利用效率高, 周转期长; 为更高效地利用好植物体内的P, P的迁移再利用率高(表 6), 因而, 衰老器官含P低, 植物通过淋溶丢失的P也少。
陈灵芝. 1983. 英国Hampstell的蕨菜草地生态系统的营养元素循环. 植物学报, 25(1): 15-23. |
冯宗炜, 陈楚莹, 王开平, 等. 1985. 亚热带杉木纯林生态系统中营养元素的积累、分配和循环的研究. 植物生态学与地植物学丛刊, 9(4): 245-255. |
侯学煜. 1982. 中国植物地理及优势植物化学成分. 北京: 科学出版社.
|
哈默特.布塞尔, 汉纳.斯可夫, 王铸豪. 1989. 华南大叶相思林的碳、氮动态的系统分析与模拟. 中国科学院华南植物研究所集刊, 4: 235-251. |
林鹏, 林光辉. 1985. 九龙江口红树林研究Ⅳ-秋茄群落的氮、磷积累和循环. 植物生态学与地植物学丛刊, 9(4): 21-29. |
卢俊培. 1994. 海南岛尖峰岭热带林凋落物研究初报. 生态学杂志, 13(1): 26-29. |
刘光崧主编.土壤理化分析与剖面描述.重庆: 科学技术文献出版社重庆分社, 1996 http://www.cnki.com.cn/Article/CJFDTotal-YYHS902.001.htm
|
拉夏埃尔(李博等译, 1985).植物生理生态学.北京: 科学出版社, 1985
|
潘开文, 刘照光. 1999a. 10年生连香树人工群落生物量研究. 应用与环境生物学报, 5(2): 121-130. |
潘开文. 1999. 四川大沟流域土壤活性铝含量及其对连香树群落的影响. 山地学报, 17(2): 147-151. DOI:10.3969/j.issn.1008-2786.1999.02.011 |
孙羲主编.农业化学.上海: 上海科学技术出版社, 1980
|
宋朝枢, 徐荣章, 张清华编. 1989. 中国珍稀频危保护植物. 北京: 中国林业出版社.
|
谭云峰, 黄建旗, 陈新媛, 等. 1989. 油茶林生态系统中营养元素循环的研究. 生态学报, 9(3): 213-219. |
王东, 高淑贞. 1990. 中国连香树科的系统研究Ⅰ.叶的宏观结构及叶柄维管束变化. 西北植物学报, 10(1): 37-41. DOI:10.3321/j.issn:1000-4025.1990.01.006 |
王东, 高淑贞. 1991. 中国连香树科的系统研究Ⅱ.次生木质部的显微和超微结构. 西北植物学报, 11(4): 287-289. DOI:10.3321/j.issn:1000-4025.1991.04.006 |
温肇穆, 梁宏温, 黎跃. 1991. 杉木成熟林乔木层营养元素生物循环的研究. 植物生态学与地植物学学报, 15(1): 36-45. |
余宁旺主编.应用数学.北京: 农业出版社, 1979
|
王夔主编.生命科学中的微量元素.北京:中国计量出版社, 1992
|
张宏陶主编.水质分析大全.重庆: 科学技术文献出版社重庆分社, 1989
|
Althur MA, Fahey JJ. 1992. Biomass and Nutrients in an Engelmann spruces subalpine fir forest in north central Colorado:pool, annuAl production, and internal cycling. Can J.For.Res., 22: 315-325. DOI:10.1139/x92-041 |
Bob Gibbons. 1995. Trees of Britain and Europe. London: Chancer press.
|
Davis G L. 1966. Systematic embryology of angiosperms. New York: Jorn wiley & Sons.
|
Erdtman G. 1966. Pollen morphology and plant taxonomy angiosperms, (corrected reprint of the edition of 1952). New York and London: Hafner publishing Co.
|
Frank B, Golley.热带森林的生产量和矿物循环.植物生态学译丛, 132~133.北京: 科学出版社, 1982, 4
|
Killingbeck K T, Costigan S A. 1988. Element resorption in a guild of understory shrub species:niche differentiation and resorption thresholds. Oikos, 53: 366-374. DOI:10.2307/3565537 |
Metcalfe C G, ChAlk L. 1957. Anatomy of the dicotyledons. Oxford, Vol.1: 30-31. |
P Duvigneaud, S Denaeyer, De Smet.陆地生态系统矿质循环.植物生态学译丛.北京: 科学出版社, 1982, 4: 144~177
|
Ratter J A, Milne C. 1973. Chromosome in Primitive Angiospermes Ⅰ, Ⅱ, notes R.B.G. Edinb, 1976: 32. |
Swamy B G L, Bailey I w. 1949. The morphology and relationships of Cercidiphyllum. J.Arnold Arb, 30: 187-210. |
Tsutsumi, T.森林生态系统中营养元素的积累和循环.植物生态学译丛, 北京: 科学出版社, 1982, 4: 177~179
|
Twilley, et al. 1986. Litter production and turnover in basin mangrove forest in southwest Florida. Ecology, 67(3): 670-683. DOI:10.2307/1937691 |