 2000, Vol. 36
2000, Vol. 36文章信息
- 高甲荣, 肖斌, 陈海滨, 刘满堂.
- Gao Jiarong, Xiao Bin, Cheng Haibin, Liu Mantang.
- 秦岭山地云杉林结构特征与更新动态的研究
- STUDY ON THE STRUCTURE CHARACTERIS TICS AND REGENERATION DYNAMICS OF CHINESE SPRUCE FORESTS IN QINLING MOUNTAINS
- 林业科学, 2000, 36(专刊1): 104-109.
- Scientia Silvae Sinicae, 2000, 36(专刊1): 104-109.
-
文章历史
- 收稿日期:1998-05-12
-
作者相关文章


2. 西北林学院 杨陵 712100
本文是在
 教授指导下完成的, 西北林学院森保89级同学参加了外业调查工作, 并得到陕西省辛家山林场的协作与支持, 西北林学院杨茂生教授、雷瑞德教授、陈存根教授等曾给予热情指导和帮助, 在此谨致谢意。
教授指导下完成的, 西北林学院森保89级同学参加了外业调查工作, 并得到陕西省辛家山林场的协作与支持, 西北林学院杨茂生教授、雷瑞德教授、陈存根教授等曾给予热情指导和帮助, 在此谨致谢意。2. Northwest Forestry University Yangling 712100
云杉(Picea asperata Mast.)林是秦岭林区高中山地带的顶极群落类型之一, 由于其蓄积量高、材质优良, 亦为秦岭林区开发利用最早的森林类型(张仰渠, 1988)。陕西省云杉天然林面积日渐稀少, 且资源分散, 因此恢复云杉林在高山地带特有的防护作用, 是秦岭林区一个重要的研究课题。本文根据对该林区天然云杉林、择伐更新林及皆伐迹地更新林的调查对比分析, 研究了云杉林的结构特征及其更新动态, 以期为探索该区云杉林的恢复途径及扩大这一树种资源提供科学依据。
1 研究区概况及方法 1.1 研究区概况该研究在陕西省宝鸡市林业局辛家山林场进行。研究区位于秦岭西部南坡, 嘉陵江上游, 34°10′~34°20′N, 106°28′~106°38′E, 海拔800~2400m。区内山峦起伏, 地势险峻, 坡度多为20°~50°, 气候属于暖温带半湿润山地气候, 在不同海拔高度, 年均温7.6℃~15.4℃, 年降水量800~1000mm, 多集中在7、8、9月。年日照时数1000~1200h, 大气相对湿度72%~86%, 年蒸发量600~800mm, 无霜期土壤为棕色森林土母岩以花岗岩为主土层厚度多为40~70cm。
1.2 研究方法在云杉天然林保留的区域韭菜坪, 选择有代表性的成熟林、择伐更新林及皆伐迹地更新林, 分别设置25m×25m的样地各1块进行调查。在样地内对胸径≥3.0cm林木进行每木检尺, 测定各林木的胸径和树高(用测高仪及目测法测定树高)。同时, 在每个样地中均匀设置9个2m×2m样方, 调查胸径 < 3.0cm幼树的数量及高度、地径、年龄和冠幅, 并调查云杉幼苗的数量、生长状况及云杉种源的数量和发芽情况等。
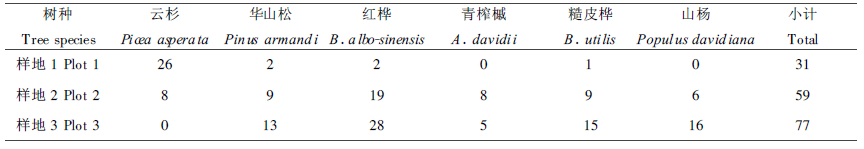
在云杉成熟林(样地1)和择伐更新林(样地2)的样地调查中, 对各主要组成树种按径级选取2~29株样木, 用生长锥分别测定各样木胸高处的年轮数, 以实测幼树至1.3m处的年龄平均值为基础确定各样木年龄。调查样地各测定树种年龄的样本数列于表 1。对云杉龄级的估算, 主要依据实测云杉样木年龄建立云杉林木年龄与胸径级的回归方程进行的。对其它树种年龄级的估算, 以实测样木年龄为参照, 根据多年林区工作经验和林木所处位置及生长状况进行目测估算的。这一方法虽不十分精确, 但可以相信, 估测年龄误差一般不会超过一个龄级。
|
|

群落组成结构是指构成群落的各种生物成分, 也是群落生长、更新和发展的基础。云杉林作为高山、亚高山地带顶极群落类型存在于秦岭山地, 占据着与其相适应的生态环境条件。云杉林的生长、发育、更新规律, 一方面决定于自身的遗传性, 另一方面在很大程度上受外界环境条件的支配(四川林业科学研究所, 1984; 李景文等, 1988)。不同生境条件下云杉林的树种组成、林分结构、生长状况和更新状况都有较大的差别。
辛家山林场韭菜坪一带的天然云杉林, 是60年代初期大面积皆伐时期保留下来的比较完整的云杉林。云杉成熟林多是以云杉为优势种构成的单优群落, 群落中常常混生有青扦(Picea wilsonii), 各林木在乔木层可以划分为两层次。主林冠层的高度为20~25m, 云杉在这一层次占绝对优势, 此外尚有少量的华山松(Pinus armandi)、青榨槭(Acer davidii)、红桦(Betula albo-sienensis)、光皮桦(Betula luminifera)等。林冠亚层为15~20m, 树种组成仍以云杉为主, 另有华山松、青榨槭、漆树(Toxicodendron verniciflum)、茶条槭(Acer ginnala)、甘肃槭(Acer mandshuricum)、鹅耳枥(Carpinus turczaninowii)等, 林下灌木主要有华北绣线菊(Spriaea fritschiana)、六道木(Abelia biflora)、鞘柄菝契(Smilax stans)、忍冬(Lonicera japonica)、紫丁香(Syringa julianae)、杜鹃(Rhododendron giraldii)等。皆伐迹地上形成的林分, 在树种组成上要比成熟的天然云杉林复杂得多。主林冠层除了华山松、红桦、光皮桦、槭树、糙皮桦(Betula utilis)外, 常常能看到山杨(Populus davidiana)、白桦(Betula platyphylla)、毛
3个调查样地中各主要组成树种在群落乔木层出现的数量见表 2。可以看出, 不同调查样地, 其主要组成树种出现的比例有较大的差异性和规律性。云杉在成熟林、择伐更新林和皆伐迹地更新林的乔木层的株数比例分别为82%、13%和0, 呈明显的下降趋势, 这与调查各样地的云杉处于不同的群落演替阶段相一致。山杨、桦木在成熟云杉林乔木层仅占总株数的10%以下, 而在皆伐迹地上则超过75%, 这也说明杨桦类为先锋树种, 在皆伐迹地有着较强的适应性和生存能力(高甲荣等, 1992)。华山松和青榨槭介于二者之间, 在不同调查样地都有或多或少的出现, 这也表明华山松和青榨槭具有一定耐荫性和生存竞争力。
|
|
群落的径级结构是指林分中各树种在不同粗度级(胸径级)上的株数分配比例, 反映了群落内不同树木个体间的竞争和分异状况。在云杉成熟林中, 云杉在各个径级几乎都有分布, 主要集中在4~8和24~36径级, 数量上亦占绝对优势(图 1), 而在其它径级中分布较少。这就表明, 在云杉成熟林中, 云杉具有很强的耐荫性, 能够在林冠下进行自我更新, 形成稳定的群落。华山松在云杉林中也有少量分布, 主要集中于20~28径级, 说明华山松具有一定的耐荫性(高甲荣, 1991)。其它树种如红桦、糙皮桦等仅在大径级中少量出现, 多系在林窗或林中空地上生长起来的先锋树种, 而在小径级中几无存在。“林窗”更新是云杉林维持其连续性的机制之一, 也是群落发生波动的重要途径(高甲荣等, 1992)。

|
图 1 云杉成熟林中各组成树种的径阶分布结构 Fig. 1 DBHgrade structure of the various tree species in the mature forest of Picea asperata |
林木的高度结构是指林分中各组成树种在不同高度级内的株数分布比例。在林木高度分布结构中, 几乎所有树种都集中于16m以上, 占调查样地林木株数总量的85%以上(图 2)。云杉在8~12m尚有少量个体存在, 这是由于云杉在庇荫条件下, 生长十分缓慢(高甲荣等, 1990)。在云杉成熟林中, 林木高生长差异尤为明显, 同一高度级的云杉, 龄级范围相差可达1~6个龄级, 而同一龄级的林木, 高度相差多达2~15m。虽然云杉幼树个体数量较少, 但却是唯一的更新树种, 这也是云杉林能够维持其群落结构相对稳定性的原因之一。而其它树种在更替层几乎没有出现, 说明云杉林下光照条件差, 抑制了其它喜光性或耐荫性较弱树种的生长, 使其难以达到主林冠层而逐渐消亡。

|
图 2 云杉成熟林中各组成树种的树高分布结构 Fig. 2 Height distribution structure of the various tree species in the mature forest of Picea asperata |
天然云杉林在进行强度择伐后, 经过了30~40年的自然生长过程, 林中的生态环境发生了变化, 与此相适应, 各主要组成树种在径级和高度分布结构上也有不同的适应性变化并呈一定的规律性(图 3、图 4)。根据各主要组成树种在择伐更新林中的径级及高度分布结构可以将其划分为4种类型: (1)种群的径级和高度分布结构相似, 主要集中分布于中、小径级和高度级, 而在大径级和高度级中数量很少或没有如云杉种群径级和高度分布结构相似主要集中分布大径级及高度级而在中小径级及高度级中极少, 如山杨。(3)种群径级和高度级分布结构相似, 在各个径级和高度级均有分布, 如红桦、糙皮桦。(4)种群径级分布与高度分布结构无一定的规律性, 如华山松、青榨槭。

|
图 3 云杉择伐更新林中各组成树种的胸径结构 Fig. 3 DBHstructure of the various tree species in the regeneraed forest under selection cutting system of Picea asperata |

|
图 4 云杉择伐更新林中各组成树种的树高结构 Fig. 4 Height structure of the various tree species in the regenerated forest under selection cutting system of Picea asperata |
可以看出, 作为耐荫性很强的云杉主要集中分布于小径阶, 而杨、桦等阔叶树种则分布于大径级和高度级。这是由于择伐后林地光照、湿度条件得以改善并拥有开阔的空间, 能满足杨、桦树种的衰退要求, 加之杨、桦等树种幼龄期生长迅速, 能很快在更新层占据优势并达到主林层。云杉早期生长速度缓慢, 在与杨桦等激烈的竞争下, 无论胸径生长或高生长都表现出小于阔叶树种(高甲荣等, 1991; 1992)。但在无外界干扰的条件下, 随着时间的推移, 云杉、华山松等最终会替代这些阔叶树而形成稳定的群落, 这是由各树种的生物学、生态学特性及其群落的环境所决定的。
2.3 群落年龄结构群落的年龄结构是指组成群落的不同树种年龄结构的综合状况, 表示不同年龄个体在树木种群中的比例关系, 也是了解林分现实状况和更新动态的一条有效途径(阳含熙等, 1988)。在云杉成熟林和择伐更新林中各组成树种的年龄分布结构如图 5和图 6所示。在云杉成熟林中, 云杉年龄差异特别明显。相差达240年, 但集中分布于幼龄(20~60年)和老龄(200~240年)阶段, 表现为多世代林木共存。对云杉胸径级与年龄级的相关规律的分析表明, 同一径级的云杉, 龄级变化范围多在1~5个龄级, 而同一龄级的云杉, 胸径级变化范围多达3~9个径级。由此可以看出, 云杉成熟林中林木径级与龄级的变化幅度较大, 这也是调查中为什么要采用目测估算法的原因之一。

|
图 5 云杉成熟林中不同组成树种的年龄结构 Fig. 5 Age structure of the various tree species in the mature forest of Picea asperata |

|
图 6 云杉择伐更新林中不同组成树种的年龄结构 Fig. 6 Age structure of the various tree species in the regenerated forest under selection cutting system of Picea asperata |
从图 5可以看出, 云杉在各个年龄阶段几乎都有分布, 但在80和180年呈间断分布。一方面, 说明云杉具有自我更新能力, 能维持群落的相对稳定; 另一方面, 反映了云杉林对外界干扰如火灾、病虫鼠害等反应的敏感性, 尚需进一步研究。而其它树种如红桦、华山松等则主要分布于60~140年, 幼龄个体几无分布, 这说明红桦等在云杉林下更新不良, 属于明显的衰退种群, 终将被云杉所取代。在云杉的生活史中, 外界生态环境的剧烈变化如火灾、风倒、皆伐等对云杉和其它树种的影响是同时存在的, 但生长初期云杉生长比较缓慢, 常常混有部分喜光性生长较快的先锋树种如山杨、桦木等, 会迅速生长占据主林层, 并能持续一定时间。同时, 生长速度中等且具有一定耐荫性的树种如华山松, 也可以部分占据主林层。这一现象正是树种生态学特性和群落生态环境变化的综合反映。
在云杉择伐更新林中, 主要组成树种个体在各年龄阶段均有分布, 但主要集中分布于30~50年之间。在对云杉林进行强度择伐后, 原有的群落生态环境发生了剧烈的变化(四川林业科学研究所, 1984), 由于不同树种的生态特性不同, 尤其是耐荫性的差异, 从而导致了在幼龄阶段各树种的年龄结构表现出明显的差异性。云杉林在强度择伐后, 林地有大量的云杉种源和幼树, 能够很快占据生境并迅速生长, 所以在30~50年的云杉林木个体中占有相当的比例。随着林木的进一步生长, 个体竞争状况趋于激烈云杉种源也随之减少所以幼龄个体也随之大为减少
从图 6可以看出, 红桦、糙皮桦、山杨种群的年龄结构非常相似, 呈明显的山峰式分布。在≥60年的老龄个体数和≤20年的幼小个体数都很小, 而在30~50年集中分布, 这是明显的衰退种群的年龄结构特征, 也与调查地的经营史相一致。
2.4 更新动态云杉林的天然更新状况, 因所处立地条件不同而有差异(张仰渠, 1988)。秦岭辛家山林区天然云杉林主要分布在平缓的沟谷和坡脚地带, 所以对不同作业方式的云杉林更新调查也仅限于这一区域。不同群落中云杉幼树、幼苗及种源状况的调查结果列于表 3。
|
|
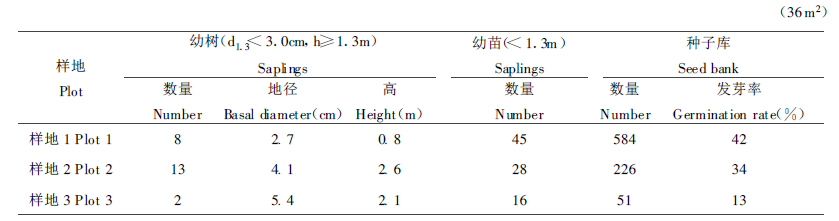
丰富的种源是云杉林实现天然更新的物质基础, 而更新幼树、幼苗的数量和生长状况则是云杉林维持稳定的决定因素。从表 3可以看出, 云杉成熟林(样地1)中存在大量的云杉种源(1.6×105个/hm2), 而且种子的发芽率也相当高, 这就为云杉林的天然更新奠定了物质基础。但是, 调查样地中云杉更新幼树的数量却相当少, 仅为2200株/hm2, 而且6~10年的幼树占40%左右, 其它树种的更新幼树不及云杉的10%, 说明成熟云杉林下的天然更新仍比较差。云杉成熟林下天然更新的特点为: (1)云杉更新幼树的分布不均匀; (2)更新幼树多呈团状或小块状分布; (3)云杉更新幼树多分布于林窗空隙或林缘; (4)云杉更新幼树数量少, 但却是唯一的更新树种; (5)更新幼树年龄分布具有近似连续性。
在秦岭林区, 对云杉林能产生剧烈影响的外界因素是火灾、风倒、病虫害和人为破坏等。根据云杉在不同群落中的年龄结构及演替动态过程, 可以认为, 稳定状态云杉林的形成至少经过3个过程, 即外界生态环境条件发生剧烈变化之后云杉幼苗幼树的侵入形成阶段(1~40年), 云杉个体之间或与其它树种之间的激烈竞争后的进展生长阶段(40~70年), 主林层的喜光性树种逐渐衰退后云杉以优势种进入主林冠层并在林下产生更新幼苗的恢复阶段(70~110年)。
根据种群对策理论, 云杉属于k对策种, 山杨、桦木等属于r对策种。由于人为采伐或火灾、病虫害等外力的强度干扰, 使原来的森林群落消失, 森林环境发生突然变化, 迹地上阳光直射气温较高, 空气湿度小表土层干燥夜晚冷却辐射增加温度急剧下降易使更新幼苗遭受日灼和霜冻的危害再加上迹地上杂草丛生, 这样严酷的生态环境使云杉难以生存。而红桦、糙皮桦、山杨等r对策种, 由于结实量大, 种子较轻易于传播, 幼苗幼树能适应温度的剧变, 而且生长迅速, 在生存竞争中有战胜杂草的能力, 因此云杉林受到强度干扰后, 桦木、山杨会立即占领新的生境, 形成桦木林或山杨林。在桦木、山杨林内, 光、热、水和土壤的水分和养分状况不断得到改善, 又为华山松、云杉等树种的种子萌发、幼苗生长提供了条件, 相反不利于r对策种群的发展。如果有足够的种子来源, k对策种群在桦木、山杨林下就可以大量更新并逐渐取代r对策种群形成混交林最终成为顶极群落类型。
3 结语云杉成熟林的树种组成以云杉为优势树种, 常伴生有少量的华山松、青榨槭和红桦。林下拥有丰富的云杉种源, 保存云杉幼树数量少且生长状况差, 但为唯一的更新树种, 因此能维持云杉种群的更新连续性和相对稳定性。
择伐更新林的树种组成以云杉、华山松、红桦等为主, 红桦等r对策种占据主林层, 而云杉k对策种处于更替层。更新林中云杉保存幼树数量多, 生长良好, 在无外界干扰的情况下将更替其它树种形成稳定的群落。
云杉林皆伐迹地上形成的林分树种组成复杂, 以r对策种占绝对优势, 云杉种源、保存的幼苗幼树数量少, 自然条件下难以恢复为云杉林或者需要较长的时间, 但在人为干预条件下可以加快和调控云杉的更新过程。
高甲荣, 王树文, 黄云鹏. 1990. 陕西省辛家山林区云杉生长状况的初步研究. 西北林学院学报, 5(1): 15-21. |
高甲荣. 1991. 秦岭林区华山松林天然更新的初步研究. 西北林学院学报, 6(1): 9-15. |
高甲荣, 刘广全, 龚立群. 1992. 秦岭火地塘林区采伐迹地天然更新的研究. 西北林学院学报, 7(2): 15-23. |
李景文, 詹鸿振, 刘传照. 1988. 小兴安岭阔叶红松林采伐迹地更新的研究. 林业科学, 24(2): 129-137. |
四川省林业科学研究所. 1984. 川西亚高山云杉林采伐迹地生态因子的变化. 林业科学, 20(2): 132-138. |
阳含熙, 伍业钢. 1988. 长白山自然保护区阔叶红松林林木种属组成、年龄结构和更新策略的研究. 林业科学, 24(1): 18-27. |
张仰渠. 1988. 陕西森林. 西安: 陕西科学技术出版社, 168-173.
|