 2000, Vol. 36
2000, Vol. 36文章信息
- 冯玉龙, 王文章, 敖红.
- Feng Yulong, Wang Wenzhang, Ao Hong.
- 长白落叶松无性系选择生理指标的研究
- STUDY ON PHYSIOLOGICAL INDICES OF CLONE SELECTION IN LARIX OLGENSIS HENRY
- 林业科学, 2000, 36(专刊1): 80-85.
- Scientia Silvae Sinicae, 2000, 36(专刊1): 80-85.
-
文章历史
- 收稿日期:1998-04-06
-
作者相关文章


2. 东北林业大学 哈尔滨 150040
2. Northeast Forestry University Harbin 150040
长白落叶松是我国东北地区森林经营和人工造林的主要树种之一,据统计仅黑龙江省每年营造的长白落叶松人工林就达6.7×104 hm2以上。为提高长白落叶松林的质量和效益,对其进行遗传改良有重要的意义。早期选择可缩短育种世代,提高选择效果,因此,一直受到国内外林业科技工作者的重视。已证明长白落叶松的早期选择是可行而有效的(杨书文等,1991;佟再康等,1990),但他们都是从表型性状直接进行选择,没有涉及与生理特性有关的研究。硝酸还原酶活力、光合速率和N、K、Zn等含量与日本落叶松生长的相关性已被证实(杨书文等,1994)。作者在研究了内外因素对长白落叶松生理特性影响(Wang Wenzhang et al.,1995;冯玉龙等,1998)基础上,研究发现硝酸还原酶活力和氮素含量(冯玉龙等,1999)、过氧化物酶等活力(冯玉龙,1996)1)和激素含量(冯玉龙等,1991)与长白落叶松不同种源的生长密切相关。本文以长白落叶松不同无性系为材料,进一步研究光合性能和氮素代谢指标与生长的关系,探讨它们用于早期选择的可行性。
1) 冯玉龙. 长白落叶松生态生理生化特性及与生长的关系. 东北林业大学博士学位论文, 1996: 90~108.
1 材料与方法把长白落叶松(Larix olgensis Henry) 9个无性系的90株一年生幼苗移栽到3个容器(50 cm×40 cm×20 cm)中,测定苗高和地径,记为H1和D01 (表 1),之后移到人工气候室(Model-E8 CMP3032,CANA-DA)中,室内条件控制为:08:00~18:00,气温25℃,相对湿度85%,光强246 μmolm-2s-1;18:00~08:00,气温17℃~25℃,相对湿度95%,无光照。5月末待针叶完全展开后,用贝克曼865型红外线CO2分析仪和ZHT型蒸腾测定仪同时测定净光合速率、呼吸速率和蒸腾速率等参数。叶室(6 cm×6 cm×12 cm)用透明有机玻璃制成,测定时采用开放气流系统,气流速度为1.5 Lm-1。叶面积用LI-3000型叶面积仪测定。不同的光强通过控制板自动调控与人工遮光等方法获得。Arnon法测定叶绿素含量及a/b比。体内法测定硝酸还原酶活力(苏梦云等,1986;周树等,1985)。比色法测定硝态氮及亚硝态氮含量(朱广廉等,1990)。纳氏比色法测定氨态氮含量(朱广廉等,1990)。所有生理指标均为两株苗木的4次测定平均值。实验结束时(1995年7月26日)测定苗高和地径,分别记为H2和D02,以H2-H1和D02-D01表示当年高和地径生长(表 1)。
|
|
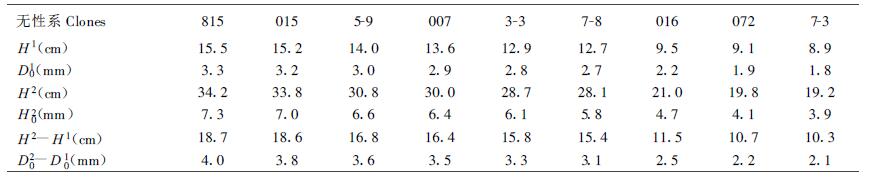
测定结果表明,长白落叶松不同无性系的光合性能变异较大(表 2)。无性系815净光合速率最高,015次之,无性系072最低,仅为815的62.8%,9个无性系的平均净光合速率为4.8267 μmolm-2s-1。无性系015的呼吸速率最高,无性系7-3最低,仅为015的27.3%,9个无性系的平均呼吸速率为1.4728 μmolm-2s-1。无性系015的真正光合速率最高,815次之,无性系7-3真正光合速率最低,仅为015的54.2%,9个无性系的平均真正光合速率为6.3017 μmolm-2s-1。方差分析表明,长白落叶松不同无性系间呼吸速率变异最大,变异系数为40.6%,真正光合速率次之,为20.4%,净光合速率变异最小,变异系数为16.6%。
|
|
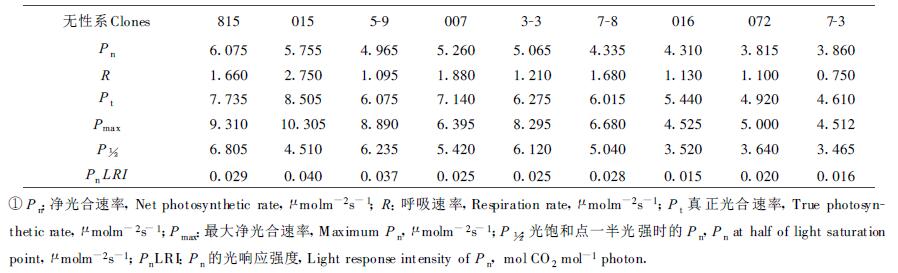
在人工气候室中测定了不同光强下长白落叶松9个无性系的净光合速率、蒸腾速率和水分利用率,分别与光强做图,并对直线部分做回归分析,导出其它有关参数,结果见表 2和图 1。不同无性系的光响应虽有差异,但基本规律是一致的,因此,图 1中只给出无性系7-3的光响应曲线。净光合速率和水分利用率与光强的线性相关范围较小,蒸腾速率与光强的线性相关范围较大,不同无性系的3个参数与光强的线性相关范围相差也较大(结果未列出)。净光合速率与光强的线性相关部分相关系数均在0.93以上,最高为0.9909,9个无性系平均为0.9518。水分利用率和蒸腾速率与光强线性相关部分的相关系数也较高,9个无性系的平均相关系数分别为0.9238和0.9152。

|
图 1 长白落叶松净光合速率、蒸腾速率和水分利用率的光响应 Fig. 1 Light response of net photosynthetic rate, transpiration rate and water utilization efficiency in L.olgensis 1.净光合速率Net photosynthetic rate; 2.蒸腾速率Transpiration rate; 3.水分利用率Water utilization efficiency |
低光强时净光合速率随光强的增加而线性升高,光强继续增加,净光合速率升高的速率逐渐变慢,最后对光强的增加不再有反应,净光合速率保持不变。由于实验时用的光强较低,在强光下时间较短,未出现光抑制现象。这与Wang Wenzhang等(1995)的测定结果相一致。长白落叶松不同无性系净光合速率的光响应强度不同,最大响应强度就是光响应曲线的初始斜率,即表观量子产量(AQY),可用来表示光合作用的光能利用效率。无性系015的最大光响应强度最高,无性系5-9和815次之,无性系016的响应强度最低,仅为无性系015的37.2%,9个无性系的平均光响应强度为0.026 mol CO2mol-1photons。
相同光强下不同无性系的净光合速率不同,光强等量增加时不同无性系净光合速率的增量也不同。光强为97.5 μmolm-2s-1时,无性系016、5-9、016和7-3的净光合速率分别为2.745、2.325、1.355和1.120 μmolm-2s-1,光强加倍(195 μmolm-2s-1)时,它们的净光合速率分别为6.595、5.900、2.790和2.610 μmolm-2s-1,分别增加了3.850、3.575、1.430和1.500 μmolm-2s-1。光强由97.5 μmolm-2s-1加倍时,响应强度大的无性系015和5-9的净光合速率增量约为响应强度小的无性系016和7-3的2.6倍。
不同无性系最大净光合速率变异较大(变异系数为30.8%),无性系015的最大净光合速率最高,无性系815次之,无性系016和7-3的最大净光合速率最低,分别为无性系015和815的43.7%和48.4%,9个无性系的平均最大净光合速率为7.102 μmolm-2s-1。为进一步说明光强与净光合速率的关系,引入了光强为光饱和点一半时的净光合速率(P½)这一参数(表 2)。无性系815的P½最高,无性系7-3的P½最低,为无性系815的50.1%,9个无性系的平均P½为4.9728 μmolm-2s-1。
从表 2可以看出,长白落叶松不同无性系的净光合速率、最大净光合速率和P½不同,它们的大小顺序也并不完全相同,净光合速率高的无性系最大净光合速率和P½并不一定高,反之亦然。这说明不同无性系的光响应方式和强度不同,净光合速率的光响应较复杂,从早期鉴定来看有必要引入多个参数。
2.1.2 长白落叶松不同无性系光合性能与生长的关系自然界中光的强度不断变化,群落内部更是如此。光合器官不可能处于恒定光强环境中,净光合速率随光强的变化而变化。因此,用3个光强下的净光合速率,即最大净光合速率、净光合速率和光强为光饱和点一半时的净光合速率,来分析解释树木的生长更合理。王文章等(1994)认为净光合速率与长白落叶松生长呈正相关,其它种落叶松净光合速率与生长的正相关性也有报道(杨书文等,1994;Ledig et al.,1974;Matysek et al.,1987a;1987b)。综合分析3个光强下的净光合速率,长白落叶松9个无性系可被分成3类,无性系815和015为最好的一类,无性系016、072和7-3为最差的一类,无性系5-9、007、3-3和7-8居中。
以表 2中的6个光合性能指标为变量,对长白落叶松9个无性系进行系统聚类分析,结果见图 2。如以10为阈值,根据这6个参数可把9个无性系分成4类,无性系016、072和7-3为一类,无性系5-9、007、3-3和7-8为一类,无性系815和015各为一类。以表 1中的6个生长性状指标为变量,对长白落叶松9个无性系进行系统聚类分析,结果见图 3。依据光合性能指标和依据生长性状指标对长白落叶松9个无性系的分类结果基本一致,表明它们之间具有内在联系。

|
图 2 长白落叶松光合性能指标系统聚类图 Fig. 2 Chart of system clustering analysis for indices of photosynthetic properties in L.olgensis |

|
图 3 长白落叶松生长性状指标系统聚类图 Fig. 3 Chart of system clustering analysis for indices of growth characters in L.olgensis |
比较分析表 1和表 2中数据可以发现,净光合速率、呼吸速率、真正光合速率、最大净光合速率、光强为光饱和点一半时的光合速率和净光合速率的光响应强度等6个光合性能指标高的无性系,6个生长性状指标亦高,如无性系815和015,反之亦然,如无性系072和7-3。表明光合性能指标与长白落叶松不同无性系的生长密切相关,回归分析结果表明它们均与生长呈正相关(表 3)。其中净光合速率与各生长性状指标的相关系数最高,最大净光合速率和真正光合速率与生长的相关系数也都在0.9以上,呼吸速率与生长的相关系数最低,但也在0.7左右。
|
|
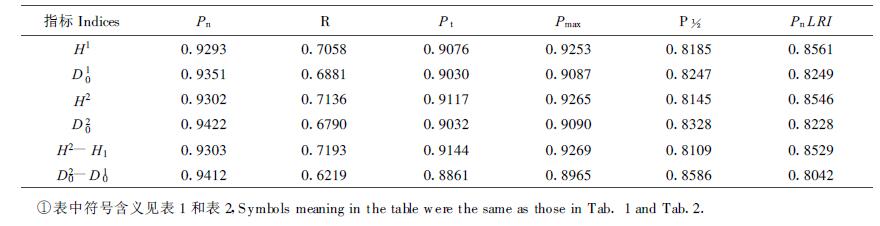
长白落叶松不同无性系光合性能的差异是受遗传控制的,因为它们生长的环境条件完全一致,这是利用光合性能指标进行无性系早期选择的基础。其它光合性能指标也有用于无性系选择的可能性,如利用光合作用的CO2响应可以筛选出适于CO2施肥和节水栽培的无性系(冯玉龙等,1999)。本文筛选出6个光合性能参数,以它们为指标把9个无性系分成4类,与以生长性状指标的分类结果基本一致,相关分析结果亦表明这6个参数与长白落叶松生长呈正相关。这与许忠志(1995)1)的研究结果相一致,他亦证明无性系815最好,无性系016和072较好,无性系007居中。
1) 许忠志. 长白落叶松扦插繁殖综合配套技术的研究. 东北林业大学硕士学位论文, 1995: 22~23
2.2 长白落叶松不同无性系氮素代谢及与生长的关系 2.2.1 长白落叶松不同无性系氮素代谢对长白落叶松不同无性系有关氮素代谢参数测定结果(表 4)表明,无性系5-9叶绿素含量最高,815次之,无性系7-3叶绿素含量最低,分别为无性系5-9和815的85%和88%。不同无性系间叶绿素a/b比差异不大,在1.81~1.85之间。不同无性系间硝酸还原酶活力变异较大(变异系数为42.1%),无性系815酶活力最高,无性系7-3酶活力最低,仅为815的38%。长白落叶松针叶硝态氮、亚硝态氮含量很低,最高不超过1.5 μgg-1,氨态氮含量较高,占总氮素(前三者之和)的绝大部分,这与对长白落叶松不同种源针叶的测定结果相同(冯玉龙等,1999)。无性系815的氨态氮及总氮含量最高,无性系7-3氨态氮及总氮含量最低,分别为815的75.1%和75.6%。总的看来,硝酸还原酶活力高的无性系,叶中氨态氮、总氮及叶绿素含量也高,净光合速率也高(表 2);硝酸还原酶活力低的无性系叶中氨态氮、总氮及叶绿素含量也低,净光合速率亦低。
|
|
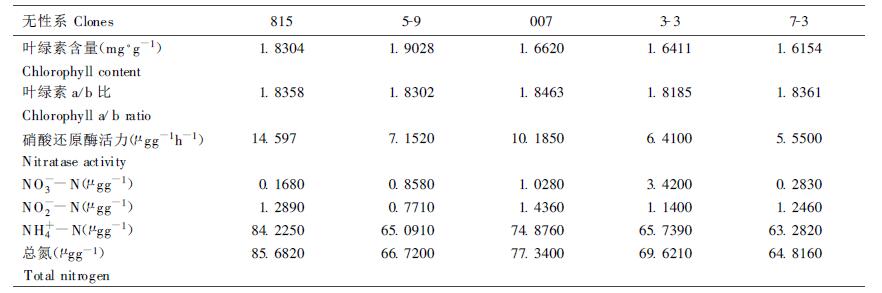
比较分析表 4和表 1中的数据,可以发现,硝酸还原酶活力强、氨态氮、总氮和叶绿素含量高的无性系生长快,硝酸还原酶活力弱、氨态氮、总氮和叶绿素含量低的无性系生长慢,氮素代谢指标与长白落叶松不同无性系的生长呈正相关。硝酸还原酶是氮素同化过程中的关键,其活力强,氮同化快,叶中可利用的氮素多,生理活动旺盛,生长快(冯玉龙等,1999)。回归分析结果(表 5)表明,硝酸还原酶活力与长白落叶松不同无性系生长性状指标的相关系数最高,叶绿素含量与生长性状指标的相关系数最低。
|
|

长白落叶松不同无性系光合性能指标变异较大,在测定的众多参数中筛选出6个与生长密切相关的指标,即净光合速率、呼吸速率、真正光合速率、最大净光合速率、光强为光饱和点一半时的净光合速率、净光合速率的光响应强度。以这6个指标为变量系统聚类分析,把9个无性系分成4类,无性系016、072和7-3为一类,无性系7-8、3-3、007和5-9为一类,无性系815和015各为一类。这与以6个生长性状指标为变量的分类结果基本一致,回归分析亦表明这6个光合性能指标与长白落叶松生长呈正相关,其中净光合速率与生长的相关系数最高,呼吸速率与生长的相关系数最低。
长白落叶松不同无性系氮素代谢指标亦不同,其中硝酸还原酶活力、氨态氮、总氮和叶绿素含量与长白落叶松生长呈正相关。回归分析表明,硝酸还原酶活力与生长的相关性最高,叶绿素含量与生长的相关性最低。
在利用生理指标进行无性系选择时,最好多个指标同时利用,或选择几个与生长相关性较大的指标不适用与生长相关性较小的少数指标。
冯玉龙, 敖红, 王文章, 等. 1998. 长白落叶松净光合速率和呼吸速率的日变化及时空影响. 河北大学学报(自然科学版), 18(2): 183-188. |
冯玉龙, 王文章, 敖红. 1999. 长白落叶松氮素营养及与生长的关系. 植物研究, 19(4): 428-434. |
冯玉龙, 王丽华, 王文章, 等. 1998. 长白落叶松激素含量与生长的关系及在早期选择中的应用. 植物研究, 18(4): 461-467. |
冯玉龙, 王丽华, 敖红, 等. 1999. 长白落叶松生理生态特性的CO2响应及意义. 植物研究, 19(1): 53-59. |
苏梦云, 周国璋. 1986. 树木组织中硝酸还原酶测定方法. 林业科技通讯, (7): 25-27. |
佟再康, 常顺国, 艾正明, 等. 1990. 长白落叶松早期选择研究. 东北林业大学学报, 18(5): 7-13. |
王文章, 陈杰, 张宝有, 等. 1994. 落叶松光合特性与初级生产力. 东北林业大学学报, 22(4): 15-21. |
杨书文, 杨传平, 夏德安, 等. 1991. 帽儿山地区长白落叶松种源选择的研究. 东北林业大学学报, 19(育种专刊): 38-46. |
杨书文, 王秋玉, 夏德安, 等. 1994. 落叶松的遗传改良. 哈尔滨: 东北林业大学出版社, 318-338.
|
周树, 郑相穆. 1985. 硝酸还原酶体内分析方法的探讨. 植物生理学通讯, (1): 47-49. |
朱广廉, 钟诲文, 张爱琴. 1990. 植物生理学实验. 北京: 北京大学出版社.
|
Ledig F T, Botkin D B. 1974. Photosynthetic CO2-uptake and the distribution of photosynthate as related to growth of larch and sycamore progenies. Silvae Genet, 23: 188-192. |
Matysek K, Schulze E D. 1987. Heterosis in hybrid larch. Ⅰ. The role of leaf characteristics. Trees, 1: 219-224. DOI:10.1007/BF01816819 |
Matysek K, Schulze E D. 1987. Heterosis in hybrid larch. Ⅱ Growth characteristics. Trees, 1: 225-231. DOI:10.1007/BF01816820 |
Wang Wenzhang, Chen Jie, Feng Yulong, et al. 1995. Study on physiological characteristics of Larix olgensis Henry and Fraxinus manshurica under controlled conditions. J Northeast for Univ., 6(1): 1-5. |