 2000, Vol. 36
2000, Vol. 36文章信息
- 马常耕, 周天相, 徐金良.
- Ma Changgeng, Zhou Tianxiang, Xu Jinliang.
- 杉木无性系生长的遗传控制和早期选择初探
- A PRELIMINARY STUDY ON GENETIC CONTROL OF GROWTH TRAITS AND EARLY SELECTION OF CHINESE FIR (CUNNINGHAMIA LANCEOLATA HOOK) CLONES
- 林业科学, 2000, 36(专刊1): 62-69.
- Scientia Silvae Sinicae, 2000, 36(专刊1): 62-69.
-
文章历史
- 收稿日期:1998-07-20
-
作者相关文章


2. 浙江省开化林场 开化 324300
2. Kaihua Forest Farm of Zhejiang Province Kaihua 3234300
林木育种学从五十年代诞生之日起,就开始了早期选择可行性的探索,因为与世代间距短的农作物相比,早期选择对林木育种事业的经济意义更大。近20年在理论和方法上都取得重大进展,早期选择正成为林木育种体系的一个有机组成部分。研究既肯定了早期选择的可行性,也看到在选择性状和适宜年龄上有树种的特异性(陈益泰,1994;Lambeth,1980;1983;Kang,1985)。杉木极易扦插繁殖,解放前我国80%左右的栽培区有直接扦插造林的习惯。因此,我们从80年代初开展杉木无性系选育研究,营造了不同树龄的各类试验林143块。本文介绍了根据树龄11和10年的两个无性系复选试验林历年树高和胸径测得的数值,对杉木无性系早期选择的可行性和方法作了初步探讨。
1 材料和方法 1.1 试验材料和田间设计两试验林都用初选的无性系建成,第1个试验包括18个无性系,用初级种子园和广西融水种源实生苗为对照。随机完全区组设计,单行6株小区,5次重复,1987年春用1 a生苗造林,土壤肥力中等。试验2包括17个无性系,用初级种子园和广西那坡种源实生苗为对照,1988年春造林,单行6~8株小区,5次重复,1、2和3重复相邻设在东向坡,土壤肥力较高,4和5重复位于西北坡,土壤肥力低。栽植行株距均为2 m×1.7 m,造林后连续抚育3 a,保存率均在95%以上。
1.2 数据收集试验1:造林当年秋开始对树高、第3 a秋开始对胸径进行连年测量,直到1997年9月为止,仅第6年未做测量。试验2:从造林后第2 a秋开始和第3 a秋起至1997年9月中旬止连年测量树高和胸径。并按国家标准材积表计算两试验林各无性系平均单株立木材积,作是否入选的依据。
1.3 统计分析内容按林木育种研究中通用公式计算了树高和胸径的年—年遗传相关、表型相关、秩次相关,历年的遗传方差和环境方差,遗传及表型变异系数,遗传力,以及历年树高和胸径与1997年最后一次测量计算出的单株材积的表型相关和遗传相关,以揭示性状自身和不同生长性状间的相关程度,为确定适宜早期选择性状和适宜树龄提供依据。同时按Mckeand (1988)的公式计算各年选择效率,以利确定最适宜早期选择的年龄段。
2 结果和分析 2.1 1997年最后一次测量时两试验林生长的平均表现树龄和树体大小极大地影响着研究结果的可靠性和实用性。与已发表的杉木无性系选择试验相较(唐基乡,1990;张全仁,1992),本研究已属最高树龄(11和10 a),它们的平均表现如表 1。
|
|

从表中数值看出,在最后一次测量时,两试验林的树体已达小径级商品材标准,方差分析结果也表明无性系间生长差异显著(P < 0.001),所以根据本试验中所测生长数据进行早期选择研究是适宜的。
2.2 遗传方差分量、遗传变异系数和遗传力的时间趋势掌握无性系遗传方差分量占总表型方差中的比率,无性系生长的遗传变异系数和遗传力等参数随树龄提高的时间趋势,对确定适宜早期选择年龄和估算选择效果有重要作用,表 2给出估算的结果。
|
|
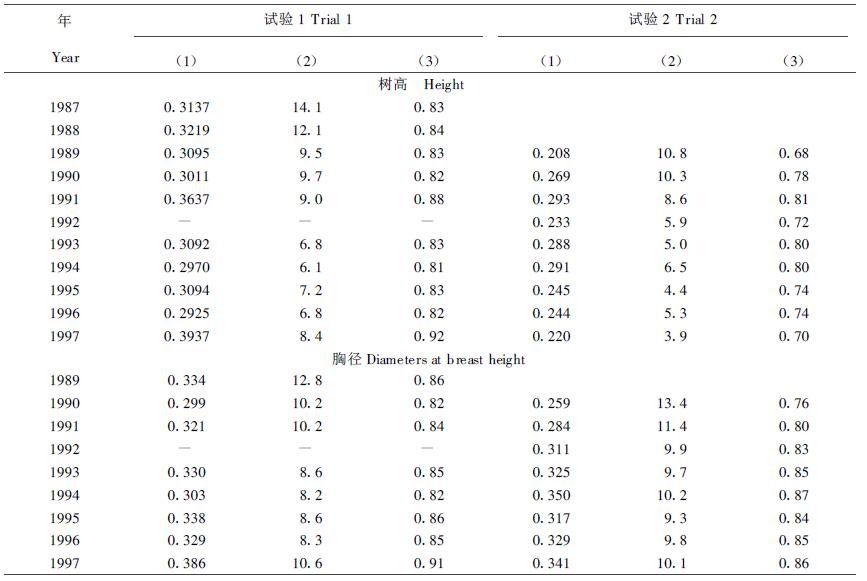
从表中数值看出,试验1的树高和胸径遗传方差分量要比试验2的高些,遗传力也表现相似趋向,这与试验2的不同重复所处立地差异较大有关。在造林4~5 a后树高和胸径的遗传方差分量和遗传力都相对达到最高,此后基本稳定在一个水平。但由于受杉梢小卷蛾危害和无性系间竞争影响,树高的遗传方差分量和遗传力波动幅度较大,并有降低的趋势。与遗传方差和遗传力的时间趋势相反,无性系间表型和遗传变异系数则随树龄增高而缩小,且树高比胸径缩减率更高。这表明,既不能用不同年龄生长差异百分率的大小变化来否定早期选择的效果,也不能用幼龄时计算出的遗传增益来阐述高龄时的选择效果,那将有夸大遗传增益的危险。
从表 2还看到,杉木无性系早期生长性状的遗传力(或重复力)极高,而胸径比树高又高些。这都与Mullin等(1994)估算的黑云杉10 a生无性系树高遗传力为0.868;Lambeth等(1994)在460个3 a生巨桉无性系上估算的树高遗传力为0.79,胸径为0.88;何贵平等(1997)对4 a生杉木无性系估算的树高重复力为0.79,胸径为0.78的结果十分接近。从而证明无性系的早期选择效果比个体选择要好。因无性系测验中的遗传力普遍比子代测验中所见个体遗传力高得多。
2.3 树高及胸径的年—年相关性状年—年相关强度是判断早期选择是否可行的重要参数。由于计算出的两个试验的结果极其相近,表 3提供试验1的结果。
|
|
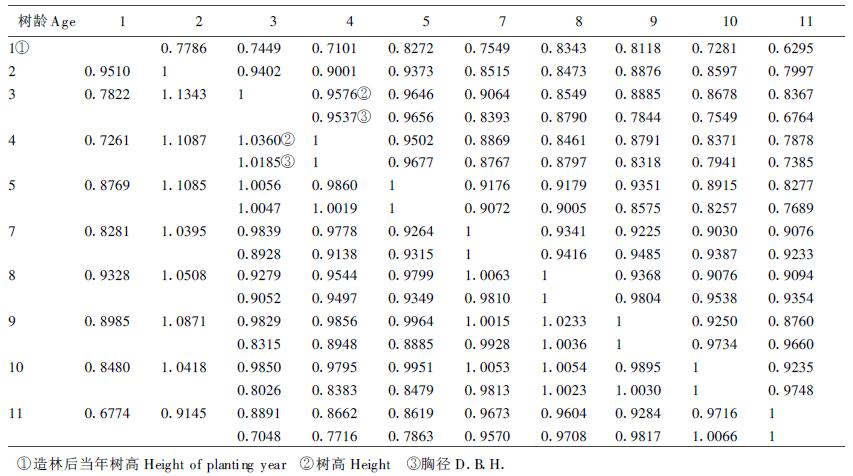
很明显,杉木无性系树高和胸径生长的年—年相关十分强,造林当年秋的幼树高与11 a生时树高的表型相关系数已达0.6295,造林3 a秋的树高与11 a生时的树高达r=0.8367。胸径相应为r=0.6764。遗传相关和表型相关的强度趋势一致,而且相关系数值要大于表型的值,证明生长的年—年相关主要由参试无性系的遗传性所决定。这种极早期就出现的生长性状年—年显著相关表明对包括杉木在内的早期速生树种的早期选择年龄会更早些。
2.4 历年树高、胸径与后期单株材积的相关提高轮伐期的单位面积材积是林木育种追求的根本目标之一,揭示造林后不同年份树高和胸径生长与采伐期材积的相关关系(本研究可假定1997年为工艺成熟的轮伐期),对性状的选用在何年开始早期选择有决定性意义。表 4给出两个试验林历年树高和胸径生长量与1997年(10和11 a生)最后一次测量时计算出的单株材积间的相关系数(秩次的和遗传的)。
|
|

从表中看出,除试验2的树高外,在造林后第3~4 a树高及胸径与后期单株材积的相关已达极显著水平,而且遗传相关系数都比按表型计算出的秩次相关要高。而令人更感兴趣的是在相同林龄时胸径与后期单株材积的相关都高于树高同单株材积的相关,表明根据胸径进行选择效果会比按树高选择更可靠些。这与近年在火炬松(Bailian,1996)、桉树(Lambeth,1994)、泡桐(Kang,1985)、日本落叶松(丁振芳,1997)早期选择研究中得出的结论一致。
2.5 无性系的生长型和生长模式由于一个基因型(个体)的无性系化增多了该基因型表型重现的次数,就为确切判断其遗传特性以更大准确性。在观察中我们看到杉木无性系的高、径生长不是等速的,而有多种配合,最明显的是除生长速度匀称配合的基因型外,还有细高的、有矮粗的,我们称谓生长型。另外也看到在试验期内不同无性系生长速度因林龄而有变化,多数为相对恒定,少数为增速型或减速型。我们称之谓生长动态模式。为有利于早期选择,对参试的35个无性系作了生长型和生长动态模式的分类,结果如表 5。
|
|
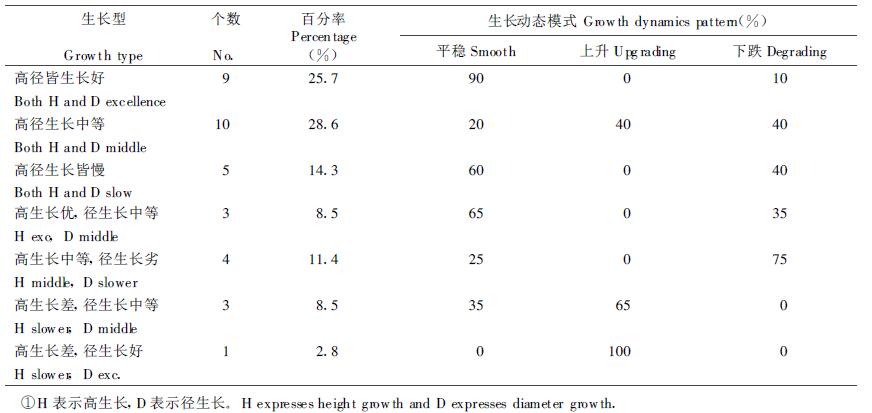
可以看出,林木的高、径生长速度的配合形式是多样的,即树高和胸径的生长力似乎受不同基因组所控制。在35个无性系中,高、径生长速度一致的无性系占68%,不一致的占1/3左右。从生长动态模式看,又是恒速型的占多数。从历年生长排序位次看,高、径生长皆快和皆慢的无性系又多属恒定型。高、径生长中速的无性系中有1/2属上升型,1/2是下跌型。高生长慢径生长快的无性系也表现为上升型。Isik等(1995)在40个欧洲云杉无性系研究中也看到过与我们在杉木中的类似现象,即存在少数细高型和矮粗型无性系,多数无性系试验期内生长位次相对恒定,也有少数早期生长慢后期加快和早期生长快后期生长转慢的无性系。其中高、径生长同速的无性系占绝大多数,两者的相关系数为r=0.83***。最令人感兴趣的是在试验中,单株材积排序居前10名的无性系中有9个表现为树高和胸径生长双优,另一个是树高生长差而胸径生长突出的,在生长双优的9个无性系中又有8个属恒定生长模式的,这对理解早期选择可行性的机制是有用的。
3 讨论 3.1 胸径指标比树高更有利于早期选择立木材积主要受树高和胸径支配,而且在高龄时胸径对材积的贡献比树高更明显。本研究中看到胸径的遗传力、年——年的相关系数与后期单株材积的相关都比同龄树高的大些,而且随林龄增高无性系间树高差异相对缩小,因而杉木无性系的早期选择以胸径为依据比树高更为可靠。这一认识可从计算出的树龄和胸径的间接选择效率来证实(表 6)。
|
|
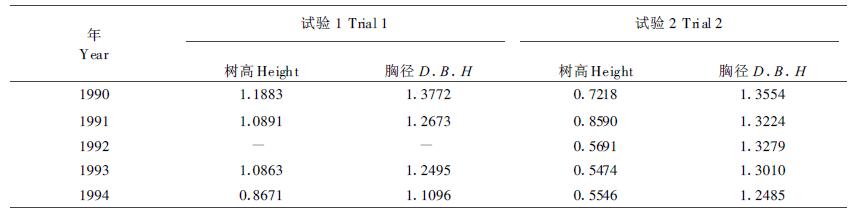
此外,由于随着树体生长,对树高的测量越来越困难,而测量胸径却容易得多,所以现在不少林木育种者都推崇按胸径进行早期选择。
3.2 早期选择的适宜树龄我们认为林木育种中讨论的早期选择概念应理解为一个年龄段,而不是某一绝对树龄。一些研究看到以树体大小作为早期选择的标志比按树龄更有意义,因树龄相同环境不同时树体大小差异可能很大。本研究看到,造林的头2~3 a,由于造林作业的冲击,幼树还不能真正发挥其遗传固有生长潜力,表现为遗传相关和表型相关值相似。而在造林后4~6 a,表型表现中的环境成分影响已经减小,此时不仅遗传力高于1~3 a时,计算出的遗传相关也比同龄的表型相关系数高,多数接近于1.0。尽管从计算出的间接选择效率值看,造林后头2~3 a的要比4~6 a生时高些(表 6),但由于无性系的生长尚处在大幅度波动期,此期选择的误选率和漏选率均较高(表 7)。综合诸多因素,4~6 a时进行早期选择是适宜的,延滞选择年龄一般不能更多地提高选择的正确率,却增加了试验林管护和测量的费用。
|
|

为实际验证早期选择的适宜性状和树龄,我们作不同树龄时按树高和胸径排序处于前1~5位的无性系与1997年按单株材积处于前1~5位无性系间吻合性比较,作为早期选择正确率的指标。同时也计算不同树龄时淘汰排序处于后1/4和1/2无性系,在1997年按单株材积表现能升入前1~5位的比率,做为早期选择时漏选的指标(表 7)。为减少篇幅,把两个试验合并计算。
从表中清楚地看到,按胸径选择要比按树高选择可靠些。如1~3 a生树高居前1~5位的无性系到后期仍优良者,因试验林不同变动在20%~80%之间,此期表现不良列入被淘汰之列,后期表现较好的达40%。说明按1~3 a树高选择和淘汰误差较大。按4~6 a表现选择时,选对率升至60%~80%,淘汰的错误率下降到10%~20%。7 a以后选择正确率和淘汰错误率没有改善。而根据胸径选择和淘汰,1~3 a时,选择正确率达80%,淘汰错误率不超过20%,在4~6 a生时选择,选对率为60%~80%,淘汰错误率仍为20%。7 a以后选择,选对率在80%~100%,淘汰错误率降至10%。有必要说明的是,出现误选和淘汰错误的原因是由于个别特异生长型无性系的干扰,如试验2中的1号无性系树高生长排序始终居后位,但粗生长却极突出,因而按树高选择属淘汰之列,最终却由于胸径生长量大,单株材积高而被选中。试验1的3号无性系表现为树高生长始终居前位,而胸径生长很差,故按树高早期就被选中,按胸径选却又被淘汰。总的看,早期淘汰的正确率比早期选优的正确率高,因早期被淘汰的无性系在以后极少能升入前1/4的位次行列,早期选中的个别无性系后期却可能跌入中等组,尽管不可能生长特别不良。
3.4 杉木无性系早期测验和选择的基本策略由于多数无性系的生长模式在试验期内是稳定的,并不因树龄而波动,早期树高和胸径生长都快的无性系在后期入选机率高;树高生长差,胸径生长快的无性系也可能中选,而树高生长快、粗生长慢的因单株材积小多不能中选。因此,杉木无性系的早期选择最好采用树高和胸径生长的双指标法,即在1~3 a生按树高初选一次,4~6 a生再按它们的胸径淘汰一次的两阶段选择策略。前一阶段按树高入选的无性系可扩大繁殖,先行利用,到6 a生时只把胸径大、单株材积居前的无性系加速扩大繁殖,其余者不再繁殖利用。因即使此期被误选的无性系,到后期表现也基本属优良一类,与最优良的无性系比,产量并没有显著差异。至于早期生长不良的无性系的淘汰,在2~3 a时进行已相当可靠,因个别被错误淘汰的无性系后期也多数不能进入优良无性系群内,最多属中等表现。而这种淘汰却减少了将进入更精细无性系试验的无性系数目,从而节省试验成本,提高试验精确性。
鉴于根据4~6 a生表现已可选出树高和胸径生长均优的无性系,所以单纯从比较选择考虑,杉木无性系比较林的行株距可由现在采用的生产性造林标准缩减至1 m或1.5 m,保证在试验地面积不增加情况下,通过包纳更多参试无性系达到提高选择效果的目的。
陈益泰. 1994. 林木早期选择研究进展. 林业科学研究, 7(专刊): 13-22. |
丁振芳, 王景章, 等. 1997. 日本落叶松早期选择技术. 东北林业大学学报, 25(3): 65-67. |
何贵平, 陈益泰, 等. 1997. 杉木无性系生长及分枝习性的遗传变异. 林业科学研究, 10(5): 556-559. DOI:10.3321/j.issn:1001-1498.1997.05.019 |
马浩, 李佩健, 等. 1998. 毛泡桐种源育种值早期预测的研究. 林业科学研究, 11(2): 179-185. DOI:10.3321/j.issn:1001-1498.1998.02.012 |
唐基乡, 邱祁生, 等. 1990. 杉木速生优良无性系选育6年研究报告. 湖南林业科技, (1): 3-7. |
张全仁, 陈佛寿等. 杉木无性系选育及其效果研究. 沈熙环主编, 种子园技术. 北京: 科学技术出版社, 1992, 306~312
|
Bailian Li, Steven E Mckeand. Genetic parameter estimates and selection efficiency for the loblolly pine breeding in the South-eastern U. S. in M. J. Matheson et al. (eds): Tree improvement for sustainable tropical forestry. Proc. QFRI-IUFRO conf., caloumdra, Queensland, Australia, 1996, 164~168
|
Isik K J. Kleinshmit, et al. 1995. Survival, growth trends and genetic gains in 17-year old Picea abies clones at seven test sites. Silvae Gen., 44(2-3): 116-128. |
Kang H. 1985. Jnvenil selection in tree breeding: Some mathematical models. Silvae Gen, 34: 75-64. |
Lambeth C C. 1980. Juvenile-mature correlations in Pinaceae and implications for early selection. For. Sci., 26: 571-580. |
Lambeth C C, Van Buijtenen J P, et al. 1983. Early selection is effective in 20-year-old genetic tests of loblolly pine. Silvae Gen., 32: 210-215. |
Lambeth C C, Endo M., et al. 1994. Genetic analysis of 16 clonal trials of Eucalyptus grandis and comparisons with seedlings checks. For. Sci., 40: 397-411. |
Mckeand S E. 1998. Optimum age for family selection for growth in genetic tests of loblolly pine. For. Sci., 34: 400-411. |
Mullin J J, Y S Park. 1994. Genetic parameters and age-age correlations in clonally replicated test of black spruce after 10 years. Can. J. For. Res., 24: 2330-2341. DOI:10.1139/x94-301 |