 2000, Vol. 36
2000, Vol. 36文章信息
- 姚庆渭, 王木林.
- Yao Qingwei, Wang Muling.
- 树木营养器官在分类上的研究与应用
- RESEARCH AND APPLICATION OF TREE VEGETATIVE ORGANS TO TREE CLASSIFICATION
- 林业科学, 2000, 36(专刊1): 10-18.
- Scientia Silvae Sinicae, 2000, 36(专刊1): 10-18.
-
文章历史
- 收稿日期:1998-07-06
-
作者相关文章


2. 中国林业科学研究院生态保护研究所 北京 100091
2. Research Institute of Forest Ecology, Environment and protection, CAF Beijing 100091
应用营养器官进行树木分类是一种理想和新的尝试,因为传统的植物分类主要根据是生殖器官,以被子植物的花为依据分类尤为明显。但因树木的花期短,季节性强,且要到一定年龄才能开花结果,造成分类工作很大困难,而树木的营养器官生长期长。常绿树种常年有叶,易于采到,如果能找出运用营养器官进行分类的途径,必然对树木分类工作带来莫大方便,而且增强其实用性。
我们观察树木营养器官通常局限于部分特征,下列内容常被忽视或不够深入,如:(1)叶脉式样;(2)髓的质地和结构;(3)叶维管束类型(叶迹、叶隙);(4)冬芽和叶痕的形态和构造;(5)枝、叶排列形式和变态。实则蕨类植物的分类,即离不开叶片脉序,古生物学家对古植物化石的叶形和脉序式样即作为分科属的重要依据。Eames,Melvill,Wanger等研究表明叶脉序式样与被子植物起源密切相关,不但有分类价值,而且对研究系统发育有明显指示作用。树木营养器官的维管系统,据Sinnott,Marsden,Horward,Forster,Arber,Bailey等的研究,同样表明其在分类和系统发育上有一定的重要性。
作者多年来,从树木冬态入手,对所有落叶及常绿树种的形态和维管束解剖作了广泛的研究,试图达到营养器官分类的目的。据初步统计我国有野生及外来栽培树木约212科1400余属近1×104种(包括树蕨,裸子植物,木本双子叶植物及单子叶植物)。现将研究所得,归纳和分述如下。
2 树木营养器官的分类依据 2.1 生活型树木是木本植物的总称,有乔木、灌木(含亚灌木)、木质藤本(或攀援灌木)之分,乔灌木约1140余属,占85%,藤本220余属,占15%。多数为陆生,极少数为附生及寄生灌木,约40属。
2.2 枝条及分枝式(任宪威等,1990)枝条有长短枝之分,树木中具有典型而稳定的短枝者约有13科14属,如银杏属Ginkgo、落叶松属Larix、海檀木属Ximenia。分枝式有二歧及多歧分枝(如槲寄生属Viscum)、单轴分枝、合轴分枝(这两种分枝式普遍存在)。另有伞形合轴分枝式,形似复伞形分枝,仅见于海桐属Pittosporum、厚皮香属Ternstroemia、杜鹃属Rhododendron、吊钟花属Enkianthus、结香属Edgworthia、马醉木属Pieris等6属,有分属价值。
枝条上有无棱角、条纹、皮孔,因属种也有一定规律,如绣线菊属Spiraea、北极果属Arctous、猬实属Kolkwitzia等属树木在1年生枝上未见皮孔。1、2年生枝色泽有的保持绿色者有卫矛属Euonymus、油桐属Vernicia、沙针属Osyris等30余属,幼枝白条者如白菊木属Leucomeris。
2.3 枝髓的结构和质地(图 1)

|
图 1 芽、髓、叶迹及叶隙节 Fig. 1 Diagram of bud, pith, leaf trace and leaf gap
芽Bud: 1.闽楠Phoebe bournei, 2.厚皮香Ternstroemia gynnanthera, 3.水冬瓜Adina racemosa, 4.香果树Emmenopterys henyi, 5.山苍子Litsea cubeba, 6.连翘Forsythia suspensa, 7.银杏Ginkgo biloba, 8.喜树Camptotheca acuminata, 9.竹节树Carallia bracthiata, 10.北五味子Schisandra chinensis, 11.黑胡桃Juglans nigra, 12.清风藤Sabia japonica, 13.领春木Euptelea pleiospernum, 14.青刚栎Cyclobalonopsis glauca, 15.二球悬铃木Plalanus acerifolia, 16.木Aralia chinensis. 髓Pith: 14.均质髓Homogenous pith, 16.海绵质髓Spongy pith, 8.薄膜髓Diaphragmed pith, 11.分隔髓Chambered pith, 6.空心髓Hollow pith. 叶迹Bundle trace: 1~6.单叶迹Simple bundle trace, 7.双叶迹Two bundle traces 8~11,三叶迹Three bundle traces, 12~16多叶迹Multiple bundle traces(叶迹5至多个,或5至多组,排列规则或不规则)叶隙节Trace gap node: 11~21.单叶隙节Unilacunar node, 11.单迹单叶隙节Simple trace from each gap(1~6), 12.双迹单叶隙节Two traces from each gap(7), 19.三迹单叶隙节Three traces from each gap (10), 20.五迹单叶隙节Five traces from each gap (12), 21.七迹单叶隙节Seven traces from each gap (13), 22~23.三叶隙节Trilacunar node, 22.单迹三叶隙节Simple trace from each gap (8、9、11), 23.多迹单叶隙节More traces from ach gap (14), 24.多叶隙节Multilacunar node,单迹多叶隙节Simple trace from each gap (15、16).(姚庆渭绘). |
双子叶树木的髓位于枝条的中心,按结构和质地有5种髓型(任宪威等,1990)。
2.3.1 均质髓髓心连续、坚硬、均质。我国主要者有76属,如壳斗科、柿科各属。
2.3.2 海绵质髓髓质松软,海绵质。我国主要有147属,如杨柳科、五加科各属。
2.3.3 薄膜髓髓质松软,中有片状横隔膜,我国有6属,如蓝果树属Nyssa、喜树属Camptotheca。
2.3.4 分隔髓髓有空室的横隔膜。我国主要有10属,如交让木属Daphniphyllum、鼠刺属Itea、杜仲属Eucommia。
2.3.5 空心髓枝条节间有空心腔。我国主要有53属,如泡桐属Paulownia、八宝树属Duabanga。
髓型变化是由于髓发育过程中破裂或形成片状隔膜所致。估计我国双子叶树木中,海绵质髓占55%,均质髓占28%,空心髓占14%,分隔髓占2.8%,薄膜髓占0.2%。
2.4 维管系统(姚庆渭,1979;任宪威等,1990;Radford AF, 1974;Howard RA, 1974)(图 1) 2.4.1 叶隙指茎维管系统向外通往叶柄的薄膜组织。(1)单叶隙节:有单迹单叶隙节(我国约有72科,如樟科、茶科、木犀科、夹竹桃),双迹单叶隙节(如银杏属Ginkgo),多迹单叶隙节(我国约有4科28属,如清风藤属Sabia、领春木属Euptelea)之分;(2)三叶隙节:我国约有77科,有单迹三叶隙节(如杨柳科、榆科)、多迹三叶隙节(如银桦属Grevillea、栎属Quercus)之分;(3)多叶隙节:我国约有22科,如木兰科、五加科、悬铃木科。
2.4.2 叶迹(1) 单叶迹:叶迹1个,我国约有61科,加茶科、杜鹃科、茜草科;(2)双叶迹:叶迹2个,仅见于银杏属;(3)三叶迹:叶迹3个或3组,我国约有77科,如胡桃科、鼠李科;(4)多叶迹:由5至多个散生或组成5至多组,我国约有63科,如壳斗科、锦葵科、梧桐科。以上3种叶迹在我国双子叶树种中约各占31%、37%、及32%接近各占1/3。
2.4.3 叶束或称顶叶迹,是叶柄顶端横切面上的维管痕,其与叶迹明显不同,有多种变化,可作分类参考。
2.5 叶片脉序(图 2、3)

|
图 2 脉序(1~20) Fig. 2 Nervation (1~20) 树蕨状二叉分枝式脉Arbuscularis: 1.毛肋桫椤Alsophila costularis,扇骨形二叉分枝式脉Flabellatus: 2.银杏Ginkgo biloba,掌状脉Palmatus, 3.直形掌状脉Reeti-palmatus:鸡爪槭Acer palmatum, 4~5.弧曲掌状脉Curvis-palmatus: 4.连香树Cercidiphyllum japonieum, 5.水青树Teteracentron sinense, 6.辐射掌状脉Radiati palmatus:粪箕笃Stephania longa, 7~8.鸟足状脉Pedatus: 7.二球悬铃木Platanus acerifolia, 8.北马兜铃Aristolochia contorta, 9~13.弧形掌状脉Convergatus: 9.药囊花Cyphotheca montana, 10.伞形寄生藤Dendrotrophe, 11.马甲子Paliurus ramosissimus, 12.耳基柏拉木Blatus ariculatus, 13.线萼金花树Blatus apricus, (14~16)掌状羽状脉Palmati-pinnate: 14.糙叶树Aphananthe aspera, 15.壳菜树Mytilaria laosensis, 16.云南紫荆Cercis yunnanensis(17~20同时参见图 3说明), 17.梯木Timonius arboreus(网状栏羽状脉), 18.肥牛木Cephalomappa sinensis(不规则梯状脉), 19.角果木Corips tagal, 20.槐花杉Amentotaxus argotaenia.(姚庆渭绘). |

|
图 3 脉序(续21~42) Fig. 3 Nervation (21~24) 羽状脉Pinnate 直羽状脉Retipinnatus:21.单一直羽状脉Craspedodrome arsitee:五桠果Dillenia indica, 22.复合直羽状脉Craspedodrome composee:大果榆Ulmus macrocarpa,弧曲羽状脉Curvipinnatus: 24.普通弧曲羽状脉Common curvipinnatus:观光木Tsoongiodendron odorum, 25.简单弧曲羽状脉Camptodromous:灯台树Borthrocaryum controversum, 26.同弓形脉Co-arcuatus:黄斑桃叶珊瑚Aucuba japonica var. variegata, 27.多弓形脉Multi-arcuatus:托叶缨Cerasus stipulacea(仿中国植物志),栅栏羽脉Femce paxillate: 28.直形栅栏羽状脉Recti-paxillate:夹竹桃Nertium indicum, 29.边缘脉栅栏羽状脉Marginal single line-continous paxillate:短序蒲桃Syzygium brachthyrsum, 30.二叉分支栅栏羽状脉Diehoromo-paxillate:齿叶赛金莲Gomphia serrata(仿中国植物志),网状栅栏羽状脉Reticulate fence paxillate(参见图版2, 17):梯木Timonius arboreus,脊形羽状脉Liratus: 31.朱蕉Cordyline fruticosa,后生羽状脉Mela-pinnatus: 32.棕竹Rhapis excelsa,二叉分枝网状脉Dichotomo-reticulatus: 33.上树南星Anadendron montana,平行脉、纵向平行脉Parallelodromous, Longirudinale: 34.粉单竹Bambusa chungii, 36.竹柏Negeia nagi, 37.大叶相思树Acacia auriculaeformis,横向羽状平行脉Parallelo-drome tranverse: 28、29、30、31,粘合羽状脉Conglutinate: 38.毛蕨属Cyclosorus,三级及较高的脉、二叉分枝网状脉3gr and higherveined, Dichotomo-reticolatus: 30、33,规则多边形脉(图 2, 6) Regular polygono-reticulatus: (Fig. 2, 6),不规则多边形脉Iregular polygonoreticulatus: (Fig. 2, 4),规则梯状脉Regular scalariformis: (Fig. 2, 9、12、13),不规则梯状脉Iregular scalariformis: (Fig. 2, 18),肥牛木Cephalomappa sinensis,方格斑纹状脉Tesselatus: 35.树形网状脉Dendro-reticulatus: 39.木犀Osmanthus fragrans,羽状网状脉Pinnate-reticulatus: 40.鼠刺Itea chinensis,汇流脉Corrivatus: 41.海芋Alocasia macrorrhiza,边缘脉Marginal veined单一延伸Simple elongate: 28.分枝延伸Branched alongate: (Fig. 2, 5),单线形连续Simple line continous: 29. (Fig. 2, 12、13),弧曲形连续Curved continous: 42.肖蒲桃Acmena acuminatissima,不完全Incomplete: 23.流苏状近缘脉Marginal limbriate: (Fig. 2, 12),珊栏羽状近缘脉Marginal paxillate: (Fig. 2, 13),叶片仅有中脉无侧脉With midribonly, without lateral veined (Fig. 2), 19.角果木Cerips tagal, 20.穗花杉Amentotaxus argotaenia.(姚庆渭绘) |
脉序亦称叶脉式样或脉型,根据Hickey, stearum, Melvillel等研究(中国科学院植物研究所,1972~1973;中国植物志编委会,1978~1998;Melvillel, 1981;1981),脉序系统有7类。
2.5.1 树蕨形及扇骨形脉二叉分枝脉,如树蕨Tree fern,银杏属Ginkgo。
2.5.2 掌状脉可分为:(1)直形掌状脉、辐射掌状脉、弧曲掌状脉,我国有21科44属,如枫香属Liquidamba、梧桐属Firmiana;(2)鸟足状脉,如悬铃木属Platanus、鞘柄木属Toricellia;(3)弧形掌状脉,我国约22科62属,如马桑属Coriaria、寄生藤属Dendrotrophe、马甲子属Paliurus、枣属Zizyphus、菝葜属Smilax。
2.5.3 羽状脉可分为:(1)直羽状脉:包括单一直羽状脉、复合直羽状脉,我国有28科57属,如榆属Ulmus 鹅耳枥属Carpinus;(2)弧曲羽状脉:包括简单弧曲形、同弓形、多弓形,我国约169科464属,如桃叶珊瑚属Aucuba、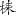
叶具多数纵向平行脉,在成对的主脉之间有第二回纵向平行脉,如竹亚科、棕榈科,在弧形掌状脉则不存在,在横向羽状平行脉中具第二回平行脉者如芭蕉科。
2.5.5 粘合羽状脉指小叶间融合的边缘线形脉,如树蕨,毛蕨属Cyclosorus。
2.5.6 三级和较高级的脉(1) 二叉分枝网状脉;(2)多边形脉:有规则多边形、不规则多边形、多角梯形;(3)梯形脉:有规则梯状、不规则梯状、方格斑纹状、梯级形;(4)小脉附属类型:有树形网状、分离羽状、羽状网状、汇流状。
2.5.7 边缘脉(1) 边缘脉缺:有单一延伸、分枝延伸;(2)边缘脉单一:有单一线形连续、弧曲形连续、不完全、不规则;(3)单一直线形边缘脉外侧有流苏状脉;(4)单一直线形边缘脉外侧有栅栏羽状脉或不规则网状边缘带。
2.6 茎、叶及其变态树木正常的叶常扁平而较宽阔,有单叶复叶之分,据统计,我国单叶树木约有1000余属,约占树木属总数的79%,复叶树木约有300余属,占21%,单叶的属中互生者约700属,对生及轮生者约370属,单身复叶者16属。茎、叶的变态,不外有肉质化、退化、刺化数种。多肉质树木我国约有20科56属,茎或叶肥厚肉质者,如芦荟Aloe、鸡蛋花属Plumeria。退化者叶退化成鳞形、卷须形或全部无叶等,我国约25属,如梭梭属Haloxylon、藤属Entada、荆豆属Ulex,刺化者有枝刺、托叶刺、皮刺、卷须刺、芽鳞刺及叶柄刺。
2.7 芽及叶痕、托叶痕(图 1)芽体因性质、部位、外形而异(任宪威等,1990)。有一型、二型之分,多数为一型,少数有花芽及叶芽二型,如木姜子属Litsea。裸芽芽体有的松散,有的拳卷紧包。芽鳞有叶身芽鳞及托叶芽鳞之分,芽鳞按数量、排列而异,通常脱落,亦有少数常年宿存于枝上者,我国约11属,如永瓣藤属Monimopetalum、藤山柳属Clematocletha。托叶芽鳞我国约有6科20属,如榕属Ficus、木兰属magnolia。叶柄脱落后枝上留有叶痕,托叶脱落后留有托叶痕,两者有时均呈环状痕者我国约29属,如白颜树属Gironniera、喜林芋属Philodendron。
2.8 附属物及内含物质(中国科学院植物研究所,1972~1973) 2.8.1 内含物质树木体内含乳状液汁者我国有11科105属,如夹竹桃科、萝科,含黄色胶汁者有山竹子属Garcinia。
2.8.2 叶含腺体等附属物包括透明油点及不透明腺体、腺鳞、腺斑、疣点、颗粒状突出、钟乳体、钙质垩点等,我国有13科64属,如白树属Gelonium 胡颓子属Elaeagnus。
2.8.3 枝叶的特殊被毛包括星状毛、簇状毛、丁字形毛、绵毛、蛛丝状毛、刚毛、刺毛,如赤杨叶属Alniphyllam、木属Cornus、紫菀木属Asterthamus。
此项附属物与含有物和植物的生理、生化关系密切,是内分泌与激素的产物。
3 树林营养器官类型的划分与分类树木在植物界虽有各个分类层次之分,但在营养器官上常有相似,难以区分,如裸子植物的松、杉、柏等科针形、刺形、鳞形叶与双子叶植物的岗松Baechea、岩高兰属Empetrum、滨樗属Suriana等极为相似,又如裸子植物的竹柏属Nagiera、贝壳杉属Agathis与单子叶植物的假叶树属Ruscus,双子叶植物的寄生藤属Dendrotrophe、金合欢属Acacia(叶状柄)、白千层属Melaluca的叶片均为纵向平行的趋同脉,非常相似,再如裸子植物的买麻藤属Gnetum为单叶对生,弧曲羽状脉,单子叶植物的菝葜属Smilax的叶具弧形掌状脉,这些脉序在双子叶植物中大量存在,易于混淆,深入探索非常必要。
3.1 树木营养器官类型的划分我国树木营养器官可分为7个类型。
3.1.1 一回复叶型一回羽状或掌状复叶。
3.1.2 二回以上复叶型二回至多回羽状或掌状复叶。
3.1.3 单叶细小、退化至宽叶无侧脉或多肉型单叶、针形、鳞形、刺形、条形、条状披针形或宽叶(均仅有中脉、无侧脉)、退化无叶,或多肉植物(茎或叶厚肉质)。
3.1.4 单叶纵向平行脉及弧形掌状脉型单叶,叶片不分裂或掌状深裂,有纵向平行脉、扇骨形二叉分枝脉或弧形掌状趋同脉。
3.1.5 单叶掌状脉型及掌状羽状脉型单叶、叶片具直形掌状脉、弧曲掌状脉、辐射掌状脉,或叶片上部为羽状脉,基部及近基部为掌状基出脉或离基三出脉。
3.1.6 单叶或单小叶互生羽状脉型单叶或单小叶(单身复叶),互生,叶片具直羽状脉、弧曲羽状脉、栅栏羽状脉、脊形羽状脉。
3.1.7 单叶对生及轮生羽状脉型单生、对生或轮生,叶片具直羽状脉、弧曲羽状脉或栅栏羽状脉。
这些类型中,掌状羽状脉型是掌状脉与羽状脉的过渡型,弧形掌状脉是纵向平行脉与掌状脉的过渡型。叶细小、退化无叶或多肉植物多数受生态环境影响产生的变化,故各自独立或全并成同个类型。类型的划分力求简明,全面,避免混淆。
3.2 树木营养器官科属综合检索表的编制为了进行树木营养器官分类的实践,作者对各个类型作了科属(含种群)综合检索表编制工作。由于树木营养器官复杂多样,因之不可能采用传统分类建立模式种的办法作为各级分类层次(科、属等)的根据和应用。在营养器官中,同一属的树种有时出现几个种群型的事实,故不可能只用某一种群型作为代表,这是与生殖器官为主分类不同之处,与此同时,我们还编制了各科分属检索表,使之更便于运用,在检索表编制过程中,我们体会到只要对树木的分类依据能确切掌握,营养器官分类是可以实现的。
4 营养器官分类的总体认识 4.1 对植物界各个分类层次的认识 4.1.1 门及亚门树蕨具二叉分枝脉及粘合羽状脉等原始脉型;裸子植物既保留原始的双叶迹及扇骨形二叉分枝脉,又有仅存中脉或中脉及侧脉均无的叶,还出现了少数向被子植物演化的弧曲状脉;进入被子植物,原始的脉型和双叶迹完全绝迹,脉序复杂多变,在双子叶植物中以三级脉的多边形及梯形为主线,叶迹亦多样;到了单子叶植物则以纵向平行脉、脊形羽状脉及羽状平行脉为主线,在平行主脉间还常出现第二回平行脉、方格斑纹状脉、梯级形脉、二叉小枝网状脉、汇流状脉及后生羽状脉。
4.1.2 科分科有的可以脉序为主线。如棕榈科、竹亚科、龙舌兰科为纵向平行脉;野牡丹科为弧形掌状脉兼具梯级形三级脉;有的以维管系统为主线,如木犀科、茶科、樟科、茜草科为单迹单叶隙、蔷薇科、蝶形花科多为单迹三叶隙;木兰科、五加科为单迹多叶隙;有的以复叶为主线,如含羞草科、辣木科为2~3回羽状复叶。有的则较复杂,但通过以科为单位的总体归纳和分析,认识到分科也有一定规律可循。
4.1.3 属营养器官分属时应注意几个问题,一是具有属型代表的属,绝不会与其它任何科属的树种混淆,如银杏属Ginkgo具有扇形二叉分枝脉、双叶迹、扇形叶片等属型,杜仲属Eucommia具有分隔髓、单叶迹、顶芽缺、体内含有胶丝等属型都是;二是有些树种的特征虽较明显,但常误作属型代表,实则不能代表属型,如山榄科金叶树属Chrysophyllum的叶片具有栅栏羽状脉误认为是属型代表,实则具有这种脉序的树木有11科17属之多,这只是一种脉型,不能代表属型,如果加上三叶迹、互生叶无托叶等特征才能成金叶树属的属型代表;三是有些属如鼠李科的鼠李属Rhammus,叶片具有弧曲羽状脉,但叶有互生和对生,芽有裸芽、鳞芽之分,小枝无刺,有的枝顶变成刺,形成4个种群型,因之4个种群型合计,才能成为鼠李属的完整属型,不可能以任何一个种群型作为该属的属型代表。检索表编制时种群型是不可忽视的。
4.2 营养器官在系统发育上的认识 4.2.1 维管系统从叶隙、叶迹上表明种子植物存在演化关系、单叶隙中,单迹单叶隙原始于多迹单叶隙,其中双迹单叶隙更具有原始性,在三叶隙中,单迹原始于多迹,在多叶隙中,单叶原始于复叶,互生叶原始于对生及轮生叶,羽状脉原始于掌状脉,总之叶隙不是弧立的,必须与其它相关特征加以综合,才能了解相应的演化趋势和规律。
4.2.2 脉序脉序的演化规律在二级脉中弧曲羽状原始于弓形脉、直羽状脉、栅栏羽状脉与羽状平行脉有相关性,三级脉中,多边形脉原始于梯形脉,更原始于树形网状脉及羽状网状脉,二叉分枝网状脉与原始的二叉分枝脉有相关性,后生羽状脉是羽状脉中最进化的脉型。
4.3 受生态环境影响产生的变态树木受生态环境影响,如高山、寒原、荒漠、草原及盐碱地、海滩等使营养器官产生特有变态,在各个门及亚门中均有出现,有的茎叶高度肉质化,有的大量退化,有的形成高山灌丛或垫状,或淹没在海岸线以下的红树林中。这些也纳入营养器官分类之中,是和生殖器官分类为主不同之处,值得引起重视。
井上赖数. 最新园艺大辞典, 第1~12卷. 诚文堂新光社, 1968~1969
|
任宪威, 姚庆渭, 王木林. 1990. 中国落叶树木冬态. 北京: 中国林业出版社.
|
西南林学院, 云南省林业厅. 云南树木图志, 上、中、下册. 昆明: 云南科技出版社, 1988~1991
|
姚庆渭. 1979. 营养器官维管系统在树木分类上的应用. 南京林产工业学院学报, 1~2: 31-52. |
郑万钧. 中国树木志, 第1~3卷, 北京: 中国林业出版社, 1983, 1985, 1997
|
中国科学院植物研究所. 中国高等植物图鉴, 第1~5册, 北京: 科学出版社, 1972~1973
|
中国科学院植物研究所. 中国高等植物图鉴, 补编, 第1~2册. 北京: 科学出版社, 1982~1983
|
中国植物志编委会. 中国植物志, 各卷. 北京: 科学出版社, 1978~1998
|
Bailey I W. 1956. Nodal Anatomy in Retrospect. Jour. Arn. Arbor, 3: 217-287. |
Bailey L H. 1958. Hortus Second ninth printing. The Macmillan Company New York.
|
Howard R A. 1974. The stem-Node-Leaf Continum of Dicotyledoneae. Jour. Arn. Abor, 55(2): 125-181. |
Hutchinson J. The Families of Flowering Plants. 2nd, edition, Oxford of the Claredon press, 1959, Vol. 1~2
|
Hutchinson J. 1967, The Genera of Flowering plants. Oxford at the Cleredon press, 1964, Vol. Ⅲ.
|
Melvillel R. 叶片结构的术语学. 生物科学参考资料, 第15集. 北京: 科学出版社, 1981
|
Melvillel R. 叶脉式样和被子植物起源. 生物科学参考资料, 第15集. 北京: 科学出版社, 1981
|
Radford A F. Vascular Plant Systematics. 1974
|
Rehder A. Manual of Cutivated Trees and Shrubs, fifth printing. Arnold Arboretum, Harvard University, 1951
|
Sinnot E W. 1914. Investigation on Angiosperms Ⅰ. The Anatomy of the Node as an aid in Classification of Angiosperms. Amer Jour. Bot, 1: 303-322. DOI:10.1002/j.1537-2197.1914.tb09371.x |