 2000, Vol. 36
2000, Vol. 36文章信息
- 王克勤, 王斌瑞.
- Wang Keqin, Wang Binrui.
- 集水造林林分水分生产力研究
- STUDY ON THE WATER PRODUCTIVITY OF THE STANDS AFFORESTED BY WATER-HARVESTING
- 林业科学, 2000, 36(专刊1): 1-9.
- Scientia Silvae Sinicae, 2000, 36(专刊1): 1-9.
-
文章历史
- 收稿日期:1998-05-12
-
作者相关文章


2. 北京林业大学 北京 100083
2. Beijing Forestry University Beijing 100083
水资源日趋紧张已是普遍存在的全球性生态问题。世界上1/3的陆地面积、涉及到50多个国家和地区,约占世界总耕地面积43%的耕地处于干旱半干旱地区(许越先,1992)。每年由于缺水造成的粮食生产损失几乎等于其他所有环境因子灾害所造成损失的总和(卢从明等,1994)。最初,农业中对水的利用和管理只局限于灌溉农业,认为在供给农作物水分方面,应在生长的各个阶段给予充足的水分,使农作物在任何时期保持尽可能高的水势和最大的蒸腾量,以获得最高产量(雷廷武等,1992)。从1996年Philip提出完整的土壤—植物—大气连续体(SPAC)的概念(Philip, 1966),到70年代以来,水分利用效率(WUE)理论的形成(Teare,1982),使农业用水研究从单一的灌溉农业扩展到普遍性的农业生产,灌溉农业在节约大量用水的同时实现了高产;旱作农业则研究如何增加少量供水以达到显著增产的目的。
林业生产依靠唯一的降水资源,受到天然降水随机性的限制,因而,对水分利用有效性方面的研究到目前为止还不多见。事实上,严重地受人为控制的人工林目前所处的现状与水分的利用有关,我国广大干旱半干旱地区人工林普遍存在的严重生态问题———“土壤干化”(杨维西,1996),主要是没有协调好水分与林木物质生产之间的关系所致。集水造林作为营造人工林的一项新技术,在时间和空间上把有限的降水资源进行再分配,使林木生长的土壤水分条件得到很大改善,促进了林木生长(王斌瑞等,1996)。但集水造林在我国仍属于实践阶段。
对于林分密度与林分物质生产之间关系的研究在90年代以前开展的较多,并且已基本形成定论。自从依田等人(1963)提出“自然稀疏3/2法则”,其通用性在20多年来引起生态学界的重视,被认为在理解生态系统中占主导地位(Westoby,1981),是植物种群生态学中的一个最一般的原理(Long et al.,1984)。由于森林的长周期性和试验的难控制性,一些试验数据与“3/2法则”有一定的出入,因而有人提出异议甚至否定意见(Sprugel,1984;Zeide,1985)。后来经重新评价(Zeide,1987)并作了一定的改进(Weller,1987)又重新确立了其重要地位,仍然认为“3/2法则”适用于大部分树种密度与材积和生物量之间的关系,树冠直径的立方与树木重量或体积成正比,即CW3∝V(Zeide,1987);大部分的研究认为,在立地条件相同的林分中,在林分未充分郁闭以前,林分的产量随密度的增加而上升(马钦彦,1983;惠刚盈等,1989;黄家荣,1993;刘君然,1994),但这是对天然林或常规措施营造的人工林研究所得出的结果。在干旱半干旱地区采用集水造林使限制林木生长的水分因子发生了很大变化,这种结论在集水造林系统中不一定适用。本文试图从林分水分生产力角度对合理的集水造林密度进行研究。
1 试验地概况试验地设在山西省方山县峪口镇土桥沟流域,属黄河中游黄土丘陵沟壑区,地处37°36′58″N,110°02′55″E,平均海拔1200 m。年均降雨量416 mm,6~9月份的降水占全年的70%以上,干燥度1.3。年平均气温7.3℃,极端高温35.6℃ (1980-05-29),极端低温-25.3℃ (1980-01-30)。年无霜期140 d。黄绵土,pH值8.0~8.4。森林草原灌丛植被区。
2 材料及方法集水措施营造的5年生和10年生刺槐林。5年生刺槐林有1,000株·hm-2、1,250株·hm-2和1,667株·hm-2 3种密度,位于立地条件一致的同一坡面,分别表示为5Y-1号、5Y-2号和5Y-3号,各密度林分中每株林木有相同面积(2 m2)的植树带,微型集水区表面为压实拍光处理,林木生长状况如表 1所示。10年生刺槐林有840株·hm-2、1,110株·hm-2、2,220株·hm-2和3,330株·hm-2 4种密度,位于立地条件一致的同一坡面,分别表示为10Y-1号、10Y-2号、10Y-3号和10Y-4号,每株林木有1.5 m2的植树带,微型集水区表面为自然坡面,林木生长状况如表 1所示。
|
|
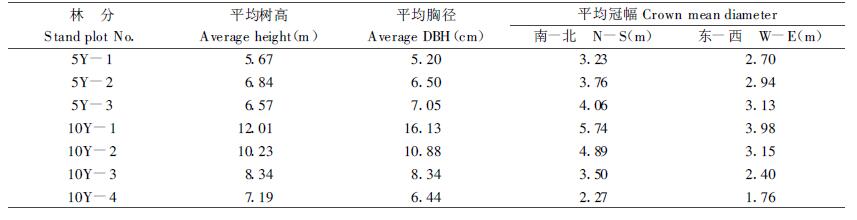
在生长季定时(每15 d 1次)定点(标准木)用LI-1600稳态气孔仪测定林木蒸腾速率,LI-3000A叶面积仪测定叶面积,гги-500型蒸发筒测定土壤蒸发量,标准雨量计观测林内外降雨量,LNW-50A型中子土壤水分仪测定土壤水分。采用每木调查法测定林木生长量,抽样烘干法获取林木地上部分生物量,标准径流场观测径流量。
3 结果分析 3.1 植树带径流汇集量(汇流量)集水造林不同于以往常规造林的关键之处在于通过修筑微型集水区人工引起径流,在植树带汇集更多的水分,缓解林木生长对水分的需求与水分不足之间的矛盾。供试的各种林分,尽管密度不同,但同一林龄的林分,其单株林木具有相同面积的植树带,因而,植树带汇流量的大小取决于微型集水区的面积和表面处理,集水区面积大、产流率高,则植树带汇流量相应也大。
根据1996年实测降雨和径流资料,计算出各林分在生长季的植树带汇流量(表 2)。结果表明,扣除树冠截留量,在5年生刺槐林试验区,集水区面积越大,植树带汇流量越大。林分5Y-1号、5Y-2号和5Y-3号的植树带汇流量分别为900.46 mm、773.05 mm和564.84 mm;同样,10年生刺槐林,以密度最小的植树带汇流量最大,即林分10Y-1号为838.59 mm,而林分10Y-3号基本接近于降雨量(462.90 mm),为501.90 mm;当密度大于3,330株·hm-2 (林分10Y-4号)时,由于冠层稠密,树冠截留率较大,植树带汇流量甚至小于降雨量,只有349.07 mm。植树带汇流量的这种大小差异将决定林地植树带土壤水分状况,进而影响到林木生长。
|
|

不论是5年生还是10年生刺槐林试验区,由于微型集水区面积林分密度的不同,使植树带汇流量随之出现了较大差异,这直接影响到植树带土壤水分。图 1(a,b)是1996年生长季5年生和10年生刺槐林试验区不同密度林分植树带土壤水分的变化曲线。表明了在同林龄不同林分密度的试验区,植树带土壤水分差异较大。林分密度越小,植树带土壤水分含量越高,年变化幅度越大。各林分土壤含水率的高峰值出现在产流集中的雨季(8月份),最小值出现在少雨的旱季(6月份)。5年生试验区中,密度最小的林分(1,000株·hm-2)植树带土壤含水率的高峰值(16.17%)比高密度林分(1,667株·hm-2)的高峰值相对高15%以上,最小值(9.41%)也偏高近12%,生长季平均偏高约10%。10年生试验区中,林分植树带土壤含水量也随密度的减小而提高。在旱季提高的幅度较小,如7月27日10Y-1号相对比10Y-4号高15.21%;雨季提高的幅度较大,8月26日10Y-1号相对比10Y-4号高33.65%。在水分亏缺的干旱半干旱区,林木生长对林地土壤水分状况的变化将会作出十分敏感的反应,最终可反映在物质生产方面。

|
图 1 两年龄组刺槐林试验区植树带土壤水分生长季变化 Fig. 1 The seasonal change curve of soil moisture in the planting strip in the experimental area of black locust forests at two different ages a. 5年生刺槐林,5-year-old locust forest; b. 10年生刺槐林,10-year-old locust forest. |
水分作为一种环境,具有双重性,既对于植物物质生产有影响,又是无机质原料用于物质生产,于是,物质生产与水分的关系极为密切。由于在植物体内所占比率最大的物质———水分绝大部分通过蒸腾作用所散失,蒸腾作用在很大程度上决定着生理活动的强弱(朗格·OL等,1985)。因而,植物物质生产与水分的关系直接表现为生产量与蒸腾量之间的密切关系。
图 2表示了5年生刺槐林试验区5Y-1号、5Y-2号和5Y-3号3种林分的林木水分生产函数曲线,不同的造林密度,单株林木年生物增量与其蒸腾量之间存在相应的函数关系。显然,3种林分中,5Y-2号和5Y-3号的水分生产函数曲线是直线,说明林木单株年生物增量随单株蒸腾量的增加直线上升,水分供应还不能使林木发挥最大的生产潜力。但曲线上每一点的斜率(年生物产量随蒸腾量的变化量dy/dx),林分5Y-2号比5Y-3号要大,证明林分5Y-3号中林木生长对水分的需求度要比林分5Y-2号大,要使林木进一步发挥生产潜力,还必须供给更多的水分。林分5Y-1号的水分生产函数曲线则是对数曲线,其初始斜率与林分5Y-2号基本相同,即在低生物量时,单株生物量随蒸腾量的加大上升较快,当年生物增量增大到一定程度(大约为10 kg·a-1左右)时,随蒸腾量上升的趋势减缓,最后基本趋于稳定,即使进一步促使林木的蒸腾作用,生物产量也不会明显增加,这时水分已不是林木生长的主要限制因子,水分供应基本能满足林木生长的需求。可见,在3种林分中,林木生长的水分亏缺度随造林密度的减小而降低。这与各林分植树带汇流量差异的大小顺序相一致,也与生长季的土壤水分高低顺序相一致,说明了水分状况决定了集水造林中林木的水分生产函数曲线。水分状况的好坏造成了不同密度林分单株林木生物量的最大值相差悬殊,5Y-1号、5Y-2号和5Y-3号的最大单株年生物产量分别为12.61 kg/株、10.69 kg/株和6.28 kg/株。

|
图 2 5年生刺槐林林木的水分生产函数曲线 Fig. 2 The function curve of water production for individual tree in 5-year-old locust forest
 5Y-1#; 5Y-1#; 5Y-2#; 5Y-2#; 5Y-3#. 5Y-3#. |
幼树生长对水分响应的敏感性较强,随着树龄的增长,敏感度会降低。但人工林在没有形成自然演替的阶段,林分稳定性较差,林木对决定性因子的反映仍十分敏感。本研究供试的林分,都表现出林木生长与水分的密切关系。5年生刺槐林如此,10年生刺槐林也表现出这一特征。4种密度的10年生刺槐林林木的水分生产函数曲线(图 3),表明了林分密度对它们的影响。密度大的林分,单株年生物产量随单株蒸腾量直线上升,即对水分需求的迫切性较强。如果能增加水分供应量,改善林地的土壤水分环境条件,以促进林木的蒸腾作用,林木单株生物产量将迅速增加。但当密度小于1,110株·hm-2时,单株蒸腾量上升到1,000 kg/株以上,单株生物产量不再随之直线上升,林木已发挥了较大的生产潜力,这时,如果再供应更充足的水分,加速林木的蒸腾作用,林木的生产能力已基本接近极限,水分生产力将会降低。

|
图 3 10年生刺槐林林木的水分生产函数曲线 Fig. 3 The function curve of water production for individual tree in 10-year-old locust forest
 10Y-1#;10Y-2#; 10Y-3#;10Y-4#. 10Y-1#;10Y-2#; 10Y-3#;10Y-4#. |
林木水分生产力表示了林木制造干物质的耗水效率,即林木消耗单位水量所生产干物质的多少。如果用WPt表示林木的水分生产力,Tr表示林木蒸腾量,E表示蒸发量,则有:
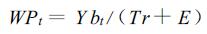
|
(1) |
其中,Ybt表示单株生物产量。林地土壤水分主要以林木蒸腾和土壤蒸发的形式所散失。由于土壤蒸发所受影响的因素多、变幅大,与林木的正常生理代谢关系不大,并可以通过各种措施进行一定程度的抑制,在林木耗水量中不予考虑,式(1)可简化为:
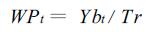
|
(2) |
图 4分别为5年生刺槐林试验区和10年生试验区中各林分的林木水分生产力随单株蒸腾量的变化曲线。表明林木的水分生产力随林分密度的降低而提高,在林木所能达到的蒸腾量范围内,高密度林分的林木水分生产力稳定不变,而在低密度(< 1,110株·hm-2)林分中,WPt在蒸腾量较低时处于较高水平,当蒸腾量达到1000 mm/株以上时,则随蒸腾量的增大而下降。这说明,在一定密度范围内,由于随着林分密度的降低,林地水分环境条件得到改善,增多了供给林木生理代谢所需要的水分,WPt随之提高。但当密度降低到一定程度后,尽管林地水分条件更为良好,但林木的生产能力已接近于最大潜力,如果再提供更多的水分,主要用于加速了林木的蒸腾作用,必然会降低水分生产力。

|
图 4 单株林木水分生产力随蒸腾量的变化曲线 Fig. 4 The change curve of water productivity varied with transpiration for individual tree at two ages a. 5年生刺槐林,5-year-old locust forest; b. 10年生刺槐林,10-year-old locust forest. |
不同林分密度造成的林木个体水分生产力之差,使各林分的平均林木水分生产力出现不同。表 3是各林分的平均林木水分生产力,5年生和10年生刺槐林中,WPt随着林分密度的减小而提高。这种大小顺序与前面所讨论的土壤含水量的变化十分吻合,进一步证实了水分与林木生长的密切关系,表现出土壤水分状况与林木水分生产之间的正相关关系。显然,在干旱半干旱地区,为了更有效合理地利用降水资源,提高水分生产力,必须有合理的造林密度。
|
|

人工采取集水造林措施的目的,是为了使林木在干旱半干旱区缺水的环境中达到正常的生长水平,从而形成良好的群落结构。由于不同造林密度对林地土壤水环境的影响存在着较大差异,使林木生长和林木个体水分生产力表现出很大差距。在供试的两林龄的林分中,以密度较小的林分林木生长状况最好,林木水分生产力最高,林木个体基本具备了“丰产理想株型”。但通过密度控制所引起的林分生态结构的不同,哪种密度的林分具有“最佳的群体”结构?还必须考虑林分的水分生产力状况。即要使林木个体和群体水分生产力实现“双优”模式标准。
如果用Ybf表示单位面积林地生物量(kg·hm-2),W表示林分供水量(mm),则林分的水分生产力可表示为:
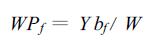
|
(3) |
表 4是1996年不同林分在生长季的林分水分生产力。1996年生长季降雨460.1 mm,在5年生刺槐林中,单株林木生物量增量最大的林分为5Y-1号(8.58 kg/株),其林分水分生产力高达18.7421 kg·hm-2·mm-1,居于首位;而密度最大的5Y-3号,两项值分别只有4.34 kg/株和15.7190 kg·hm-2·mm-1。对于10年生刺槐林试验区,4种密度中仍是以最小的林分水分生产力最高,1996年生长季林分水分生产力可达50.6703 kg·hm-2·mm-1,较大的两种密度(1,110和2,220株·hm-2)其林分水分生产力相差不显著,分别为27.4172 kg·hm-2·mm-1和29.0450 kg·hm-2·mm-1,最大密度的只有10.6179 kg·hm-2·mm-1。可见,随着林分密度的增大,由于林地水环境不断恶化,林木个体的生长发育逐渐减缓,到最大密度(3,330株·hm-2)时,林木生长基本停滞,甚至干枯死亡,群体严重分化,甚至到了难以恢复的程度。同时,对两林龄林分水分生产力的比较可以看出,随着林龄的增长,林分的合理密度也发生变化,刺槐林从5年生的幼龄林到10年生的中龄林,根据“双优”模式,林分的合理密度也从1,000株·hm-2左右下降到840株·hm-2左右。
|
|
从林分整个生育期水分生产力水平来看(表 5),在5年生刺槐林试验区,3种密度的林分水分生产力几乎相当,在8.00~8.50 kg·hm-2·mm-1之间,但从单株林木水分生产力可以看出,最小密度的林分(1000株·m-2)最高(6.7144 g·kg-1);10年生刺槐林中,仍以最小密度的林分(840株·hm-2)为理想的群落结构,其水分生产力达到21.6646 kg·hm-2·mm-1,林木水分生产力也达到12.1632 g·kg-1。根据“双优”模式,5年生和10年生刺槐林都以最小密度的林分个体和群落水分生产力最高。从造林来讲,集水造林的密度不应大于1,000株·hm-2,但小于这一密度,从林木水分生产力曲线可知,将降低林木水分生产力,也难以形成稳定的林分。
|
|
集水造林是通过造林密度的合理调节对降雨的再分配改善林木生长的土壤水分环境,使林分发挥其正常的生产潜力,因而,集水造林模式的合理与否可用水分生产力这一理论指标定量化地加以分析。
不同的集水造林模式其水分生产力出现高低的根本原因在于对林分水环境改善程度的大小。以集水区产流、植树带汇流为主要特征的集水造林对水环境的改善首先表现在植树带汇流量的大小。植树带汇流量决定于微型集水区的面积大小,即决定于集水造林的密度。随着林分密度的增大,植树带汇流量大幅度减少,进而影响到植树带土壤水分状况出现相应的变化规律。5年生刺槐林试验区,低密度林分(1,000株·hm-2),的植树带土壤水分比高密度林分(1,667株·hm-2)相对偏高15%;10年生刺槐林试验区,低密度林分的植树带土壤水分普遍偏高于高密度林分,在旱季,林分10Y-1号相对比林分10Y-4号偏高15.12%,在雨季可偏高30%以上。
植树带土壤水分状况的不同,将影响林木的物质生产,使不同密度林分的林木水分生产表现出各自的规律。林分密度从高到低,林木水分生产函数曲线由直线转为对数曲线,并且年生物量增量随蒸腾量的变化量(曲线的变化斜率dy/dx)由小逐渐增大,即随着林地水分状况的改善,林木蒸腾作用的增强,林木能进一步发挥其生产潜力。但林分密度减小到一定程度,曲线斜率(dy/dx)将由增加转而逐渐降低趋势,林木的水分生产力将开始下降,继续改善水分状况不利于水分利用效率的提高,说明集水造林从林木个体生产的角度看有其合理密度。在合理密度的范围内,林木个体水分生产力将达到较高水平。5年生最小密度的刺槐林(1,000株·hm-2)和10年生中最小密度的刺槐林(840株·hm-2),林木水分生产力分别达到6.7144 g·kg-1和12.1632 g·kg-1,居同龄所有林分之首,达到了“丰产理想株型”模式标准。
个体最优的林分在群体水平上是否合理,可用林分水分生产力这一指标加以衡量,上述的两个林分其林分水分生产力在同龄组中也属最优,分别达到8.41 kg·hm-2·mm-1和21.66 kg·hm-2·mm-1,即达到了“最佳群体结构”模式标准。
采用“双优”模式,将林分的供水状况和林木的耗水过程与林分的物质生产紧密相结合,从水分利用的角度确定集水造林的合理密度,不仅考虑到提高林木个体的生长水平,而且还考虑到林分群体结构的稳定性,既提高了林分的经济价值,也使林分能正常发挥其生态防护功能。对于干旱半干旱区造林工作,水分为主要的限制因子是一个现实性问题,从水分利用效率来考虑和确定合理的造林模式,无疑具有十分重要的理论和实践意义。
黄家荣. 1993. 贵州马尾松人工林经营密度模型初探. 北京林业大学学报, 15(4): 32-37. |
惠刚盈, 罗云伍. 1989. 江西大岗山丘陵区杉木人工林生产力的研究. 林业科学, 25(6): 584-569. |
[德]朗格·OL等, 樊梦康等译. 水分与植物生活———问题与研究现状. 第一版. 北京: 科学出版社, 1985: 310~344
|
雷廷武, 曾德超. 桃树叶水势的调控及其对生长量的影响. 见(in)许越先. 农业用水有效性研究. 第一版. 北京: 科学出版社, 1992: 81~86
|
刘君然. 1994. 林分密度理论及应用. 第一版. 北京: 中国林业出版社, 9-27.
|
卢从明, 张其德, 匡廷云. 1994. 水分胁迫对光合影响的研究进展. 植物学通讯, 11(增刊): 9-14. |
马钦彦. 1983. 华北油松人工林单株林木的生物量. 北京林业大学学报, 5(4): 1-16. |
王斌瑞, 王百田. 1996. 黄土高原径流林业. 第一版. 北京: 中国林业出版社, 103-122.
|
许越先. 1992. 农业用水有效性研究. 第一版. 北京: 科学出版社, 1-5.
|
杨维西. 1996. 试论我国北方地区人工林植被的土壤干化问题. 林业科学, 32(1): 78-85. |
Long JN , Smith FW. 1984. Relation between size and density in developing stands : A description and possible mechanisms. For. Ecol. Manage., 7: 191-206. DOI:10.1016/0378-1127(84)90067-7 |
Philip J R. 1966. Plant water relations: some physical aspects. Ann. Rev. Plant Physical, 17: 245-268. DOI:10.1146/annurev.pp.17.060166.001333 |
Sprugel DG. 1984. Density, biomass, productivity, and nutrient cycling changes during stand development in wave-regenerated balsam fir. For. Ecol. Manage., 54(1): 23-43. |
Teare I D. 1982. Crop-Water Relations. A Wiley-Inter science Publication, 1-20.
|
Weller DE. 1987. A re-evaluation of the -3/2 power rule of plant self-thinning. For. Ecol.. Manage., 57(1): 23-43. |
Westoby MandHowell J. 1981. Self-thinning: The effect of shading on glashouse populations of silver beet (Beta vulgaris). J. Ecol., 69: 359-365. DOI:10.2307/2259672 |
Yado K, Kira T, Ogawa H, et al. 1963. Self-thinning in overcrowed pure stands under cultivated and natural conditions. J. Boil. Osaka City Univ., 14: 107-109. |
Zeide B. 1985. Tolerance and self-tolerance of trees. For. Ecol.Manage., 33(2): 517-537. |
Zeide B. 1987. Analysis of the 3/2 power low of self-thinning. Forest Sci., 33(2): 517-537. |