 2000, Vol. 36
2000, Vol. 36文章信息
- 喻理飞, 朱守谦, 叶镜中, 魏鲁明, 陈正仁.
- Yu Lifei, Zhu Shouqian, Ye Jingzhong, Wei Luming, Chen Zhengren.
- 退化喀斯特森林自然恢复评价研究
- A STUDY ON EVALUATION OF NATURAL RESTORATION FOR DEGRADED KARST FOREST
- 林业科学, 2000, 36(6): 12-19.
- Scientia Silvae Sinicae, 2000, 36(6): 12-19.
-
文章历史
- 收稿日期:1999-04-26
-
作者相关文章


2. 南京林业大学 南京 210037;
3. 茂兰国家级自然保护区管理处 荔波 558400
2. Nanjing Forestry U niversity Nanjing 210037;
3. The Management of Maolan National Reserve Libo 558400
喀斯特区是一种生态环境脆弱区。喀斯特森林是一种脆弱的生态系统, 因不合理利用, 引发水土流失, 导致生境退化, 甚至形成“石漠化”, 严重威胁农业生产环境乃至人类生存。退化喀斯特森林的恢复与重建意义重大。
退化群落自然恢复实质是群落进展演替。对我国南方退化喀斯特群落研究可追溯到本世纪40年代(侯学煜, 1946;1952;郭魁士, 1940), 半个世纪以来, 对喀斯特森林顶极群落的特点、属性、退化原因、过程、生境条件及树种适应性、种群及更新特征、结构功能等研究较多, 并提出了退化群落自然恢复途径(王献溥, 1981;胡舜士, 1982;肖育檀, 1988;方任吉, 1988;徐樵利, 1993;梁士楚, 1992;朱守谦, 1987;1993;1995;1997;张家来, 1993;张祝平, 1993;杨继镐, 1990;杜道林, 1996;杨汉奎, 1991)。但对退化群落自然恢复评价缺乏研究。因此, 在阐明退化喀斯特群落自然恢复的生态学过程基础上进行恢复评价研究具有理论和实践意义。
1 研究区域概况本研究在贵州茂兰国家级自然保护区内进行。地理位置为东经107°52′~108°05′, 北纬25°09′~25°20′。区内主要出露岩石为纯质石灰岩和白云岩, 属裸露型喀斯特地貌。年均温18.3℃, ≥10℃积温5767.9℃; 全年降水量1320.5 m, 集中分布于4~10月; 年均相对湿度80%;年均霜日7.3 d; 全年日照时数1272.8 h, 日照百分率29%。属中亚热带季风湿润气候, 有利于林木生长。土壤以黑色石灰土为主, 土层浅薄且不连续, 剖面构型多为AF-D型、A-D型。地表水缺乏, 土体持水量较低, 土壤富钙和富盐基化, pH 6.5~8.0, 有机质含量高。
保护区多数地段是中亚热带原生性喀斯特森林, 为常绿落叶阔叶混交林(周政贤, 1987), 也有不同退化程度的演替群落, 对退化群落自然恢复的研究有很强的代表性。
2 研究方法本研究采用以“空间代替时间” (D.Mueller-Dombois et al., 1986)的方法, 建立退化群落自然恢复的演替系列。在分析群落组成、结构、功能变化的基础上, 提出评价退化群落自然恢复的3个指标, 即退化群落自然恢复的潜力度(restoration potentiality, RP)、恢复度(restored degree, RD)和恢复速度(restoration speed, RS)。
潜力度(RP)计算借用相似度系数公式(王伯荪, 1987)计算,

|
(1) |
式中:CS为群落更新库中幼苗库或土壤种子库的组成结构与更高演替阶段群落组成结构之间的相似度系数, 即幼苗库的潜力度(RPs1)或土壤种子库恢复的潜力度(RPs), A为幼苗库或土壤种子库各种组的株数百分率总和, B为更高演替阶段的群落组成结构各种组的株数百分率总和; C为A和B中共有种组中株数百分率低值的总和。
恢复度(RD)分两部分计算:一部分是以各群落结构功能特征指标为相似元, 采用下列计算各相似元恢复度,
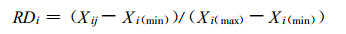
|
(2) |
式中:RDi为第i个相似元的恢复度; Xij为演替系列中第j个群落第i个相似元指标, Xi (min)为第i个相似元中的最小值, Xi (max)为最大值。
另一部分为群落组成结构恢复度, 借用(1)式计算, 式中CS为演替阶段群落与顶极群落间种组结构的相似度系数, 即群落组成恢复度, A为演替阶段群落各种组株数百分率总和, B为顶极群落种组总株数百分率总和, C为A和B中共有种组中株数百分率低值的总和。
群落各相似元恢复度(RDi)与群落年龄(A)之间的关系用逻辑斯蒂生长模型(郎奎健, 1989)进行拟合,
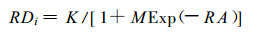
|
(3) |
其中, K为最大恢复度即1.0;M, R为参数。
恢复速度(RS)根据式(3)导出。
野外调查群落样地中每个乔灌木个体的种类、起源、胸径(或地径)、高度、盖度等。乔灌木生物量采用W=a (D2H) b的回归式推算, 草本层生物量采用刈割法实测。土壤种子库特征采用萌发试验测定。用于本研究的样地共55个, 18454 m2, 其中顶极群落固定样地2000 m2, 破坏性试验固定样地600 m2。乔灌木生物量样木149株, 萌发试验样品54份。
3 结果与分析 3.1 自然恢复阶段将54个群落样地通过样地聚集、聚集团联结、样地定位等一系列步骤, 构建退化群落自然恢复的演替系列, 并用最优分割法将退化群落自然恢复过程分为草本群落阶段、草灌群落阶段、灌丛灌木阶段、灌乔过渡阶段、乔林阶段和顶极群落阶段6个演替阶段(喻理飞, 1998)。喀斯特地区生境复杂, 能适应同一演替阶段生境的物种不是一个而是具有相同适应性的多个物种构成的种组。根据群落组成物种在自然恢复过程中优势地位的变化, 划分为如下5个适应等级种组1) :先锋种为阳性树种, 具光补偿点、饱和点高, 平均净光合速率高, 耐旱性强的特点; 次先锋种的光补偿点、饱和点较高, 平均净光合速率较高, 耐旱性较强; 过渡种的光补偿点较高, 平均净光合速率低, 具有一定的对弱光的利用能力, 耐旱性较弱; 次顶极种的光补偿点较低, 平均净光合速率较高, 耐荫性较强, 耐旱性较强; 顶极种为耐荫树种, 光补偿点低、饱和点高, 平均净光合速率中等, 耐旱性较强。
1) 喻理飞, 退化喀斯特森林自然恢复的生态学过程研究, 南京林业大学博士学位论文, 1998
自然恢复过程是由先锋种经过渡种最终为顶极种替代的发展过程(喻理飞, 1998)。各演替阶段及主要优势种为:草本群落阶段除草本植物外, 主要树种有悬钩子(Rubus palmatus Thunb.)、多花木蓝(Indigofera amblyantha Craib.)、小果蔷薇(Rosa cymosa Tratt.)、金樱子(Rosa laevigata Michx.)、全缘火棘(Pyracantha atalantioides (Hance) Li)等先锋种。草灌群落阶段除上述先锋种外, 主要有
|
|
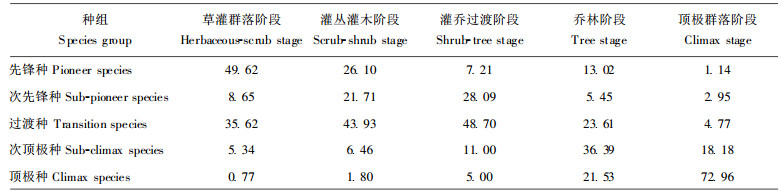
群落更替与群落更新层和主林层的组成结构相关。群落更新层中个体数量来自于群落下层的幼苗、幼树和土壤中可萌发种子数量, 因此, 群落更新库由群落下层幼苗、幼树和土壤中可萌发种子数量构成, 前者为群落幼苗库, 后者为土壤种子库。各演替阶段群落及其更新库数量特征见表 2、3。
|
|
|
|

假设自然恢复过程中一个种组被另一种组更替的概率与更新层中后一种组的数量成正比, 那么更新库种组组成结构与更高演替阶段群落种组组成结构的相似性可表征为群落自然恢复潜力, 即相似性越高, 向更高演替阶段发展的潜力越大, 反之亦然。据此, 将退化群落更新库组成结构与更高演替阶段群落组成结构间的相似度定义为退化群落自然恢复潜力度(RP)。
用(1)式分别计算群落幼苗库和土壤种子库的恢复潜力度RPs1和RPs (表 4)。由于它们对群落恢复潜力度的贡献率随演替阶段变化而异, 如土壤种子库在群落自然恢复过程早期作用大, 后期作用小, 而幼苗库不论实生或萌生, 始终起着重要的作用, 因此, 采用层次分析法(姜启源, 1993)确定RPs1和RPs权重。草本群落阶段至灌木灌丛阶段RPs1、RPs权重分别为0.7500, 0.2500;灌乔过渡阶段至顶极群落阶段分别为0.8950, 0.1050。根据RPs1和RPs及权重, 计算得出群落恢复潜力度RP (表 4)。结果表明:低一级演替阶段群落总是向相邻更高一级演替阶段群落恢复的RP最高, 退化群落自然恢复是由低级阶段向高级阶段顺序替代过程。自然恢复早期, RP较高而后期较低。这与各阶段群落组成和物种侵入特点有关。早期阶段, 群落组成以阳性先锋种占优势, 群落高度低、盖度小, 先锋种的种实小、重量轻, 易到达退化群落中, 并能适应早期群落环境, 迅速萌发生长, 故RP高。后期阶段, 群落组成以耐荫顶极种为主, 其种实大且重, 种子量小, 加之动物搬运、取食, 能萌发生长进入更新库的数量相对减少; 而阳性先锋树种仍可随风侵入, 因其幼苗具有一定耐荫性, 加之自然干扰也常形成局部透光明亮生境, 使其在更新库中总有一定比例, 导致RP下降。
|
|

退化群落自然恢复过程可视为与原顶极群落的结构、功能从低相似度向高相似度的发展过程, 其自然恢复终极是与原群落相同的植被型。据此, 将群落恢复度(RD)定义为退化群落通过自然恢复在组成、结构、功能上与顶极群落阶段的最佳群落的相似程度。
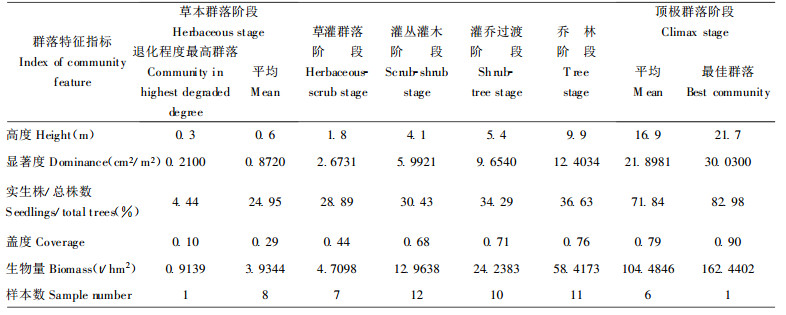
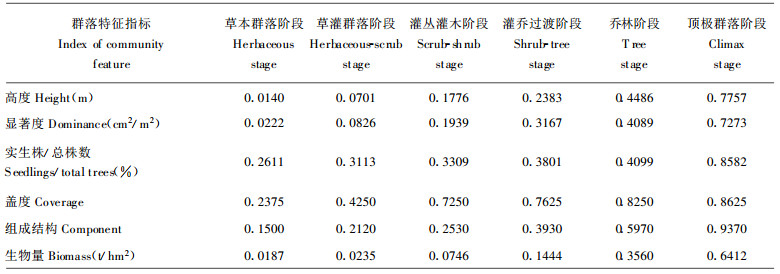
各演替阶段群落与顶极群落阶段的最佳群落之间的相似程度, 选择表征群落结构功能特征的群落高度、显著度、实生株/总株数、盖度、生物量、组成结构6个指标为相似元进行测度, 用(2)式分别计算前5个指标的恢复度, 用(1)式计算组成结构恢复度, 得各结构功能指标恢复度(表 5)。采用层次分析法确定各指标恢复度权重, 群落高度、显著度、实生株/总株数、盖度、组成结构、生物量的权重分别为0.0877、0.1753、0.0877、01234、0.2630、0.2630;根据各指标恢复度和权重, 得RD (表 6)。
|
|
|
|
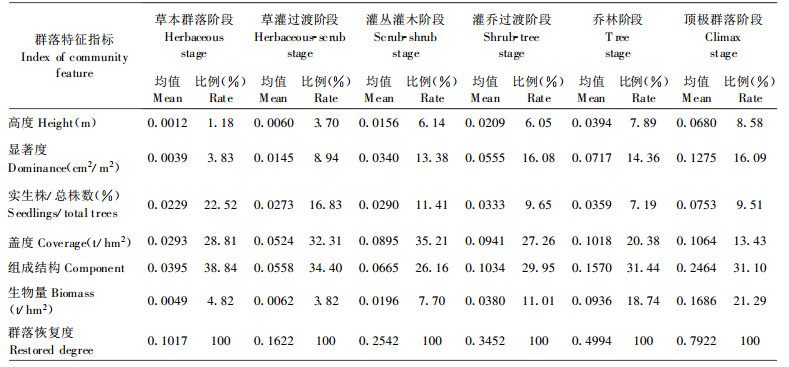
表 5、6表明:退化群落自然恢复是恢复度逐渐提高的过程。群落恢复度在灌丛灌木阶段达0.2542, 乔林阶段约达0.5, 顶极群落阶段约达0.8, 达到群落组成、结构、功能的恢复。群落各特征指标恢复度百分率反映该指标对群落恢复的贡献和重要性。在恢复过程中, 组成结构恢复始终起重要作用, 说明其调整是群落恢复的核心和实质; 盖度在早期和中期阶段作用较大, 实生株/总株数在早期阶段作用较大, 而生物量则在后期阶段作用较大。这反应出各恢复策略的变化, 早期阶段, 以更新对策为主, 萌生株/总株数的比例高约69%~85%;中期阶段, 林内光环境改变, 导致自然稀疏, 树种替代, 其恢复对策是组成结构调整; 后期阶段, 组成结构和生物量的重要性提高, 恢复对策是进一步调整组成结构、积累能量, 向结构合理、功能完善的顶极群落发展。
3.4 恢复速度自然恢复过程中, 因生境变化速度、组成物种生长发育规律和树种替代变化等差异, 使退化群落恢复并非匀速提高, 即在各阶段每提高一定的群落恢复度所需要的时间不同。将单位时间内群落恢复度(或各特征指标恢复度)向顶极群落方向发生的位移定义为群落自然恢复速度(RS)。单位时间内群落恢复度发生位移的距离越大, 群落自然恢复速度越快, 反之亦然。
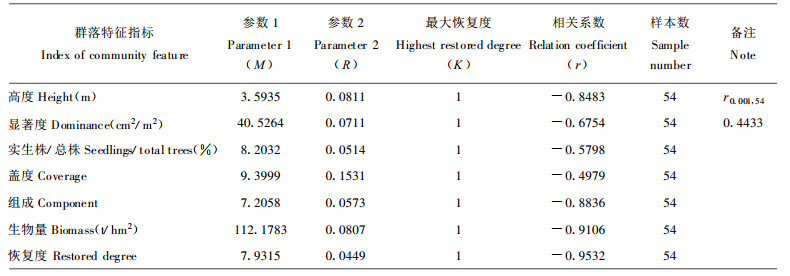
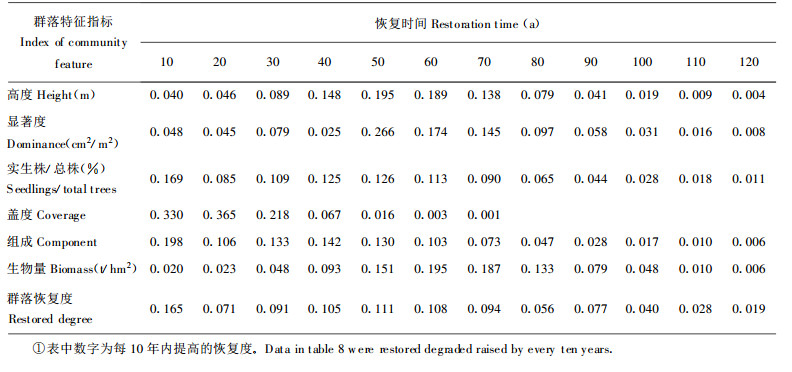
对演替系列中54个样地群落组成、结构、功能指标恢复度与群落年龄的关系用(3)式拟合, 结果见表 7。根据回归关系式, 可得表 8、9。
|
|
|
|
|
|
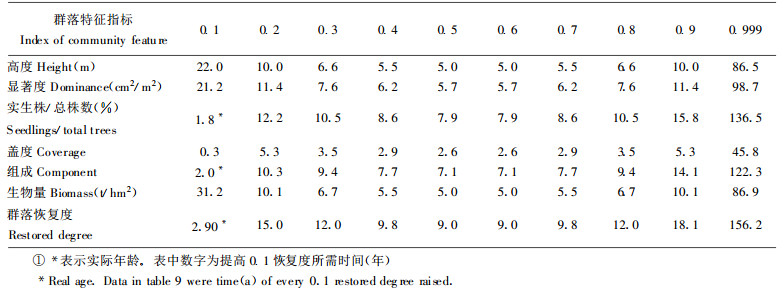
各群落特征指标恢复速度不同, 若以恢复度0.8计为基本恢复, 群落盖度、组成结构、高度、实生株/总株数、显著度的恢复时间分别需用23 a、60 a、66 a、68 a、71 a, 生物量的恢复则需用76 a, 说明群落结构恢复快于功能恢复。群落恢复度达0.8需时最长, 约80 a, 反应了群落整体功能的恢复滞后于群落结构功能各分量的恢复。群落恢复的早期, 其恢复速度较慢, 特别是恢复度从0提高到0.1时, 高度、显著度、生物量需用20~30 a, 但群落组成、盖度却费时不多, 约2 a, 原因在于退化群落所受干扰非彻底性毁灭, 总是保留有原群落植物繁殖体, 并以萌生方式迅速生长。破坏试验表明:彻底砍伐原有群落一年后, 萌生数量达4524株, 占总株数的72.15%, 盖度达0.25。中期阶段, 恢复速度快, 恢复度由0.4提高到0.7仅需用10~30年, 后期阶段, 群落恢复度高于0.8, 恢复速度慢, 尤其是恢复度从0.9提高至0.999时, 需近100年, 反应了群落结构功能完全恢复极为困难。退化群落从草本群落阶段恢复至灌丛灌木阶段需用近20 a, 至乔林阶段约需47 a, 至顶极群落阶段则需近80 a。
4 结论与讨论本研究以“空间代替时间”的研究方法建立退化群落自然恢复的演替系列, 在分析退化群落恢复过程中群落组成、结构、功能变化的基础上, 用恢复潜力度、恢复度、恢复速度三个指标评价退化喀斯特群落恢复是可行的, 有利于揭示和加深理解退化喀斯特群落自然恢复的生态学过程, 并可为南方大面积退化喀斯特群落恢复提供评价依据, 具有现实意义。在研究条件限制的情况下, 此法值得推广。当然, 若能采用长期定位研究, 会取得更满意的效果。
退化喀斯特群落自然恢复其主体是由低级阶段向高一级阶段顺序替代过程, 但因退化群落中原有群落组成成分的繁殖体存在, 使退化群落直接向更高演替阶段发展具有一定的潜力, 深入研究退化群落自然恢复的生态学过程, 有助于进一步了解演替过程的多变性和复杂性。
退化喀斯特森林自然恢复的早期阶段恢复潜力度较高、恢复度低、恢复速度较慢; 中期阶段恢复潜力度高、恢复度中等、恢复速度快; 后期阶段恢复潜力度低、恢复度高、恢复速度慢。群落整体恢复速度低于群落各特征指标恢复速度。在群落各特征指标中, 群落高度、盖度、实生株/总株数、组成结构的恢复速度高于生物量恢复速度, 即群落结构恢复速度快于功能恢复速度, 尤其是盖度和组成结构恢复最快, 因此, 退化群落从草本群落阶段开始恢复40~50年, 达到有较为正常的组成、外貌和结构的森林, 但要达到森林功能的完全恢复则需时很长。
退化喀斯特群落自然恢复过程中, 其恢复对策变化是由早期更新对策向中期结构调整对策至后期结构功能协调完善对策更替。深入研究恢复对策及其变化, 对于制定退化喀斯特森林经营措施, 加速退化喀斯特森林恢复具有重要意义。
杜道林, 等. 1996. 茂兰喀斯特山地广东松种群结构和动态初步研究. 植物生态学报, 20(2): 159-166. |
方任吉等.亚热带石灰岩山地存在常绿阔叶林—以南川金佛山为例.见钟章成编著, 常绿阔叶林生态学研究.重庆: 西南师范大学出版社, 1988 : 537~549
|
郭魁士. 1940. 广西石灰岩区土壤之初步观察. 土壤, 1(4): 32-47. |
侯学煜. 1946. 贵州盘县之植物组合与土壤之初步观察. 土壤, 5(1): 53-61. |
侯学煜. 1952. 贵州省南部植物群落. 植物学报, 1(2): 65-106. |
胡舜士, 等. 1982. 广西阳朔石灰岩山地乌冈栎的群落学特点及其在植被分类中的位置. 植物学报, 24(3): 264-272. |
姜启源. 1993. 数学模型式. 第二版. 北京: 高等教育出版社, 305-335.
|
郎奎健, 等. 1989. IBM PC系列程序集—数理统计, 调查规划经营原理. 中国: 中国林业出版社, 24-27.
|
梁士楚. 1992. 贵阳喀斯特山地云贵鹅耳枥种群动态研究. 生态学报, 12(1): 53-60. DOI:10.3321/j.issn:1000-0933.1992.01.013 |
王伯荪. 1987. 植物群落学. 北京: 高等教育出版社, 54-55.
|
王献溥, 等. 1981. 广西石灰岩地区常绿、落叶阔叶混交林的群落学特点. 东北林学院学报, (3): 30-47. |
肖育檀. 1988. 湖南石灰岩石山青冈栎落叶树混交林的群落分析. 中南林学院学报, 8(2): 146-154. |
徐樵利. 1993. 中国南方石灰岩荒山开发利用新探. 自然资源学报, 8(2): 115-121. DOI:10.3321/j.issn:1000-3037.1993.02.003 |
杨继镐, 等. 1990. 广西大青山石灰岩山地土壤理化性质的演替及其造林绿化. 林业科学, 26(5): 402-409. |
杨汉奎, 等. 1991. 贵州茂兰喀斯特森林群落生物量研究. 生态学报, 11(4): 307-312. DOI:10.3321/j.issn:1000-0933.1991.04.010 |
喻理飞, 等. 1998. 退化喀斯特群落自然恢复过程研究—自然恢复演替系列. 山地农业生物学报, 17(2): 71-77. |
张家来. 1993. 应用最优分割法划分森林群落演替阶段的研究. 植物生态学与地植物学学报, 17(3): 224-231. DOI:10.3321/j.issn:1005-264X.1993.03.006 |
张祝平, 等. 1993. 粤北石灰岩山地主要造林树种的生理生态学特性. 植物生态学与地植物学学报, 17(2): 133-142. |
周政贤主编.茂兰喀斯特森林科学考察集.贵阳: 贵州科技出版社, 1987 : 1~23
|
朱守谦等.茂兰喀斯特森林初析.见周政贤主编.茂兰喀斯特森林科学考察集.贵阳: 贵州科技出版社, 1987 : 210~224
|
朱守谦主编.喀斯特森林生态研究(Ⅰ).贵阳: 贵州科技出版社, 1993 : 1~73
|
朱守谦主编.喀斯特森林生态研究(Ⅱ).贵阳: 贵州人民出版社, 1997 : 9~47, 55~107, 128~159
|
朱守谦. 1995. 茂兰喀斯特森林生物量构成初步研究. 植物生态学报, (4): 354-367. |
D Muller-Dombois, H Ellenberg著.鲍显诚, 张绅等译.植被生态学的目的和方法.北京: 科学出版社, 1986 : 247~252
|