 2000, Vol. 36
2000, Vol. 36文章信息
- 刘济明.
- Liu Jiming.
- 茂兰喀斯特森林主要树种的繁殖更新对策
- THE REPRODUCTIVE AND REGENERATIVE COUNTERMEASURES OF THE MAIN WOODY SPECIES IN MAOLAN KARST FOREST
- 林业科学, 2000, 36(5): 114-122.
- Scientia Silvae Sinicae, 2000, 36(5): 114-122.
-
文章历史
- 收稿日期:1998-12-16
-
作者相关文章


一种森林植被类型, 一个树木种群, 在地球上怎样发展与存在下去, 树种的繁殖与更新是关键。关于植物的繁殖与更新, 国外已作过许多工作, 如“Population Biology of Plant”、“Seed Ecology”、“Introduction to Plant Population Ecology”、“Plant Reproductive Ecology”等(Harper, 1977; Michale, 1985; Silvertown, 1982; Willson, 1983)。近年来国内也作了一些研究, 如剌五加生殖生态研究(祝宁, 1992), 栲树种群结实及动态研究(刘济明, 1996; 钟章成, 1988), 红松种群研究(刘庆洪, 1988), 喜树种子扩散研究(刘济明等, 1997), 光皮桦生殖生态研究等(朱守谦, 1997)。本文从植物开花、结实、种子雨、种子库及其萌发这一过程来研究茂兰喀斯特森林部分主要树种的繁殖与更新方式, 旨在为合理利用、保护及恢复喀斯特森林植被提供一些参考。
1 研究地概况研究地位于贵州省南部黔、桂交界处的茂兰国家级自然保护区(25°09′~ 25°20′N, 107°52′~ 108° 05′E)。区内为典型的喀斯特峰丛地貌, 山峰海拔860 ~ 1010 m, 洼地海拔450 ~ 600 m。成土母岩以中下石炭纪白云岩及石灰岩为主。年均温15.3 ℃, 7月均温26.4 ℃, 1月均温8.3 ℃, ≥10 ℃积温5727.9 ℃, 年均降雨量1320.5 mm, 全年平均相对湿度83 %。土壤以黑色石灰土为主, 土层浅薄, 地面岩石裸露, 土被不连续, 土壤pH7.5 ~ 8.0, 有机质和全氮含量特别丰富(周政贤, 1987)。样地设在洞也山坡中部, 此处为南坡33°, 海拔670 m, 岩石裸露率60 %, 森林植被保存完好, 没有林窗。样地群落组成如表 1所示, 其中重要值为5个样地统计而得(样地面积900 m2, 每样地分为10个90 m2的样方), 多度采用Braun-Blanquet多度级。
|
|
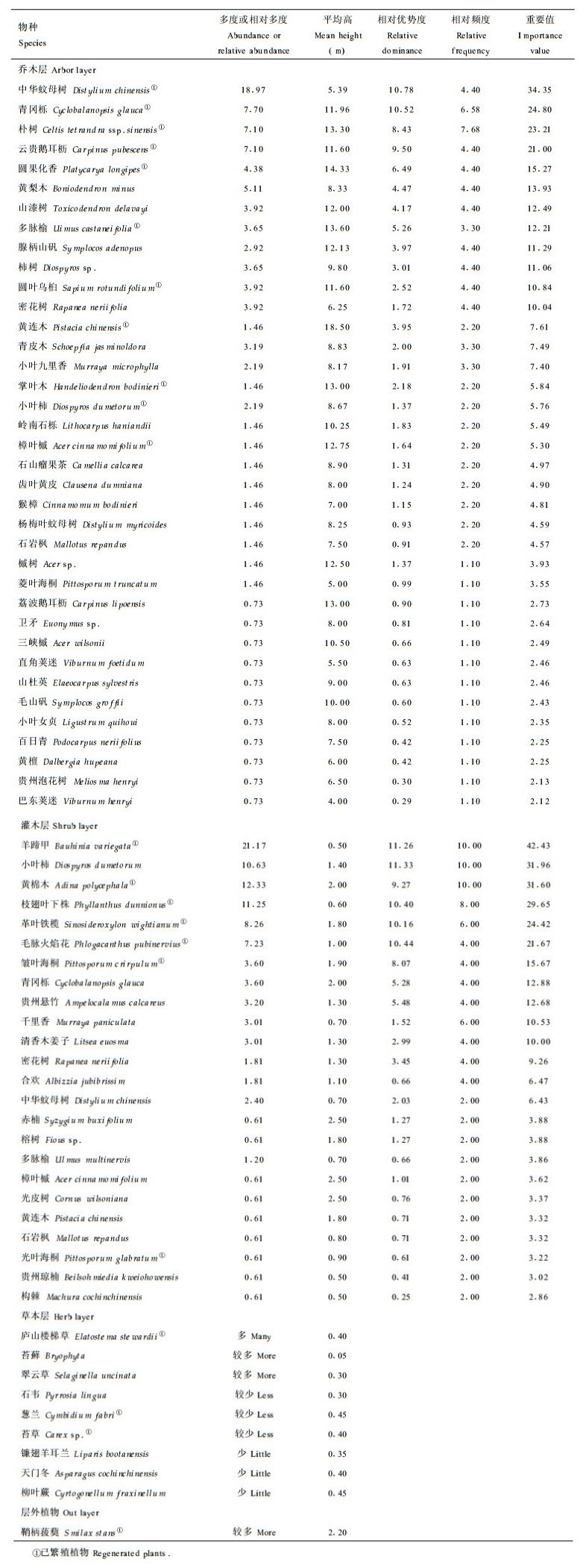
样地所在群落为喀斯特常绿落叶阔叶混交林群落, 郁闭度0.95, 其生活型谱构成中, 高位芽植物占绝对优势, 达91.79 %, 地上芽植物、地面芽植物、地下芽植物和1年生植物分别占3.49 %, 2.05 %, 1.44 %和1.23 %(朱守谦, 1993)。群落优势种不明显, 重要值分散, 垂直结构比较复杂, 层次界线不明显。群落水平结构复杂, 主要表现在植株的成丛性明显, 群落内的植株通常是成丛生长于石沟石缝之中, 植丛的大小、丛间距离及其分布极不规则。
2 研究方法在喀斯特森林保存完好的地段设置5个30 m ×30 m的固定样地, 对样地内木本植物进行群落生态学调查, 并观察样地附近林窗处幼苗幼树情况。对样地内主要组成树种的开花结实进行定位观察。采集各树种一定数量的成熟种子置于无盖玻璃瓶内, 将玻璃瓶瓶口向下埋入样地内土壤中, 每月定期取一定数量于25 ℃~ 28 ℃恒温培养箱中作发芽试验。在样地内均匀设置40个种子收集器收集种子雨。每年种子散播结束种子库天然萌发开始时的1月上旬和种子库天然萌发结束的5月下旬, 在种子收集器旁取40cm ×50cm的枯枝落叶及其下10cm深的土壤, 将种子放在枯枝落叶和土壤于适宜条件下萌发(Thompson et al., 1979), 连续观察1年, 统计出苗的种类和数量, 并以此反映库中活力种子情况。同时, 镜检枯枝落叶和土壤中植物种子情况。在每个种子收集器附近设置一个1 m ×1 m的小样方, 观察并统计种子库的天然萌发。1993 ~ 1996年连续研究4年取平均值。
3 结果与分析 3.1 主要树种的选择对样地进行群落生态学调查后得知, 在900m2样地内, 乔木层有树种37种, 灌木层有树种25种, 层外木本植物1种。结合样地实际情况, 选择已处于繁殖阶段的群落主要组成树种(乔木层中重要值前20位的树种, 灌木层中重要值前5位的树种及1种层外木本植物)为观察研究对象。它们是中华蚊母树(Distylium chinensis)、青冈栎(Cyclobalanopsis glauca)、朴树(Celtis tetrandra ssp. sinensis)、云贵鹅耳枥(Carpinus pubescens)、圆果化香(Platycarpus longipes)、圆叶乌桕(Sapium rotundifolium)、多脉榆(Ulmus castaneifolia)、黄连木(Pistacia chinensis)、小叶柿(Diospyrus dumetorum)、掌叶木(Handeliodendron bodinieri)、樟叶槭(Acer cinnamomifolium)、羊蹄甲(Bauhinia variegata)、枝翅叶下株(Phyllanthus dunnionus)、革叶铁榄(Sinosideroxylon wightianum)、鞘柄菝葜(Smilax stans)和黄棉木(Adina polycaphala)等16种。其中朴树、云贵鹅耳枥、圆果化香、樟叶槭、黄连木、黄绵木、鞘柄菝葜、圆叶乌桕和掌叶木在样地更新层中无幼苗。在林窗处, 这16种植物均有大量幼苗。
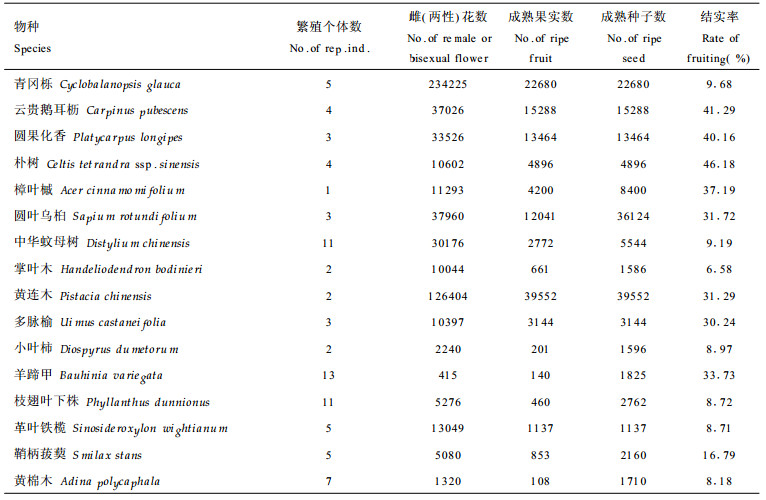
3.2 主要树种的开花、结实与种子雨(1) 开花与结实 种子植物种群在自然界的延续和发展, 大多以开花结实的形式产生后代不断补充死亡个体或扩大已有种群。样地内主要树种的开花结实情况如表 2所示。由表 2可知, 各树种结实率差异较大, 掌叶木结实率最小, 仅为6.58 %, 朴树的结实率最大, 达46.18 %。(2)种子雨 植物种子成熟之后以种子雨的形式进入种子库。林中动物对树上种子进行采食会造成成熟种子的一些损失, 一些种子被动物完全消费掉或带走, 一些种子被动物消费一部分, 但剩下部分没有发芽能力, 因而种子雨中包括能发芽的成熟有效种子和成熟被害种子两部分(这里不讨论未成熟种子)。各树种种子雨情况如表 3所示。表 3中的种子损失率为表 2中成熟种子数减去种子雨中的成熟有效种子数后与表 2中成熟种子数之比。从表 3可知, 种子损失率最小的是圆果化香(17.78 %), 最大的是青冈栎, 达53.57 %。此阶段的种子损失是由于动物的采食所至。据观察, 猴类和啮齿动物啃食、搬运及虫类的蛀食较为普遍, 它们主要危害较大的种子。鸟类损害中小型种子。
|
|
|
|
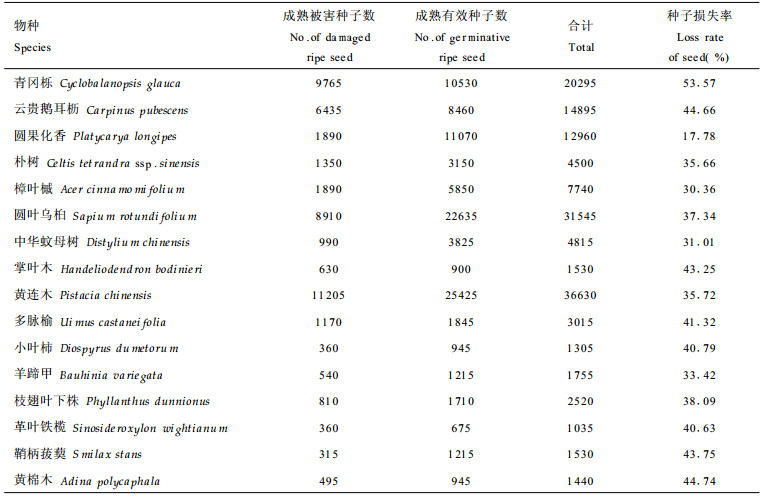
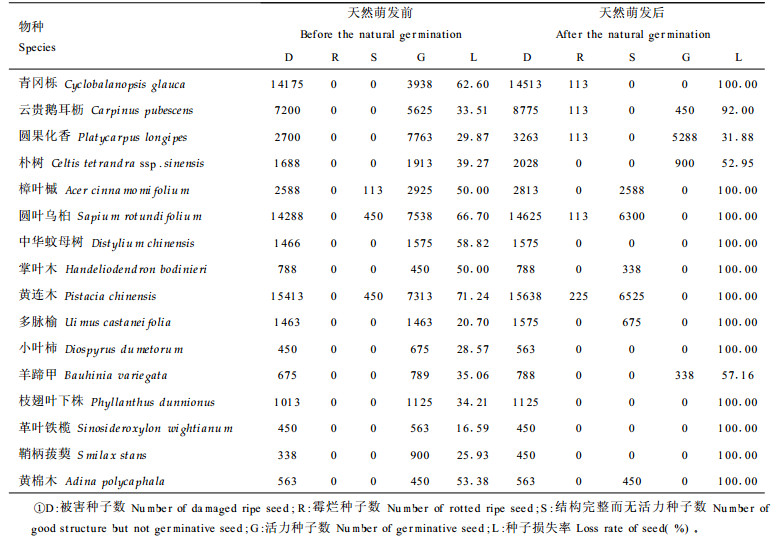
植物种子以种子雨形式进入种子库后, 在种子库中等待适宜的萌发条件。等待萌发的过程中会由于各种原因造成种子的进一步损失, 如动物采食及搬运、病菌引起霉烂变质、种子自然衰老等。通过镜检及萌发试验得表 4中种子库结构。种子进入种子库时, 最先总是位于土壤表面, 在种子散播后种子库天然萌发前这段时间, 树上种子很少, 动物就大量采食地面种子, 主要是啮齿类动物、蚁类和虫类。另外也有一些物种种子自然衰老较快, 一部分种子开始丧失活力, 因而引起种子损失, 如表 4的樟叶槭、圆叶乌桕、黄连木等, 在天然萌发前的种子库中就发现一些结构完整而无活力的种子。此间种子损失率较大, 黄连木种子的损失率最大, 达71.24 %, 损失率最小的是革叶铁榄, 仅有16.59 %。到种子库天然萌发结束时, 种子在种子库中由于各种原因进一步损失, 一部分被动物损害, 一部分被病菌侵染而霉烂变质, 一部分因自然衰老而丧失活力, 还有一部分已经萌发出苗, 种子库中所剩活力种子很少, 只有圆果化香、云贵鹅耳枥、朴树和羊蹄甲还有活种子存在, 其它种类都因各种原因活力种子全部损失。种子在种子库中的保存状况与物种的更新对策及种子的天然寿命有关。在种子成熟时采集种子埋于土壤中, 定期检查发芽率变化即可知道种子的天然寿命。以上各树种种子的发芽试验结果如表 5所示。由表 5可知, 不同树种, 其种子寿命不一样。大多数种子寿命较短, 在一个萌发季节后即丧失活力。云贵鹅耳枥种子可保持活力2 a, 朴树和羊蹄甲种子可保持活力3 a, 而圆果化香种子可保持活力4 a以上。
|
|
|
|
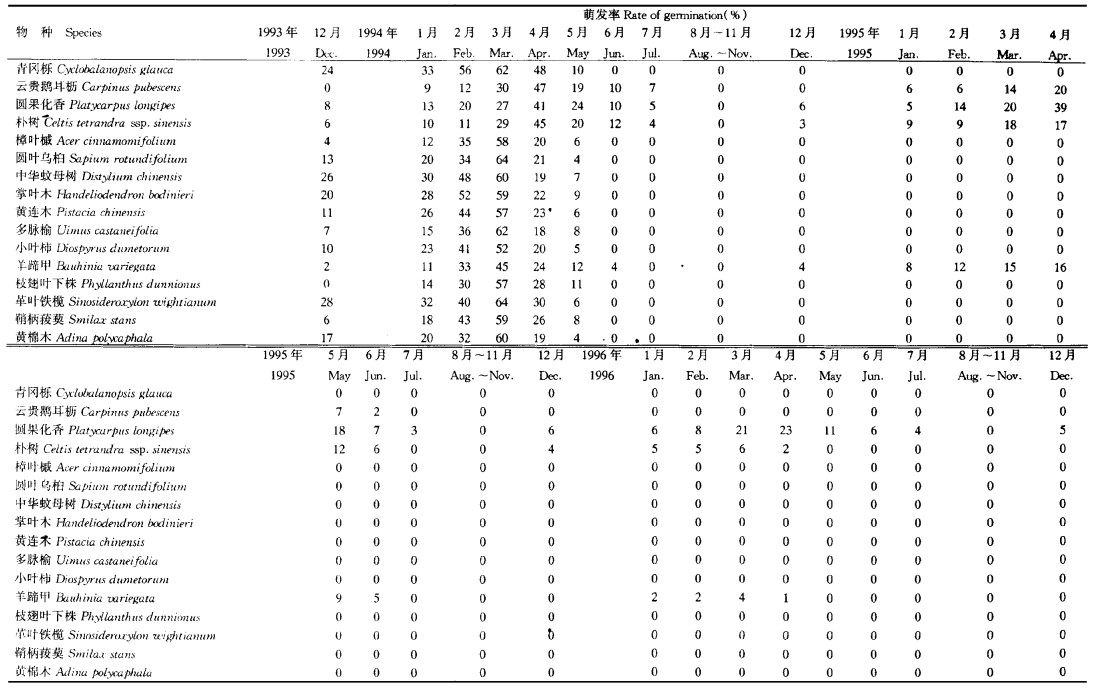
种子库中的活力种子在适宜条件下即可萌发成幼苗补充进现有种群中去, 但各物种的更新对策不尽相同, 现有群落内环境条件对库中各类种子的适宜程度也不完全一样, 因而各类种子的萌发及萌发幼苗的存活状况也会有一定差异。各主要树种库中种子的天然萌发状况如表 6所示。从表 6可知, 各树种萌发出苗及幼苗当年的存活状况差异较大, 可分为3种类型:第1种是库中活力种子没有萌发, 第2种是库中活力种子萌发出苗, 但幼苗的当年存活率为零, 第3种是库中有活力种子萌发出苗, 当年也存活一定数量的幼苗。将种子库、各树种种子发芽试验结果、库中种子天然萌发状况、群落更新层及林窗中幼苗状况进行比较分析可知, 这些树种的繁殖更新对策可分为4种类型: (1)种子成熟入土后迅速形成幼苗, 幼苗在林下缓慢生长, 以幼苗的形式等待林窗出现, 林窗出现后幼苗迅速生长补充到现有种群中去。这类树种, 种子寿命较短, 在天然萌发前的种子库中有活力种子, 萌发季节内出现幼苗, 并且当年存活一定数量, 萌发季节结束后的种子中无活力种子, 也没有结构完整而无活力(即自然衰老丧失活力)的种子。如青冈栎、中华蚊母树、小叶柿、枝翅叶下株和革叶铁榄等。(2)种子成熟入土后迅速形成幼苗, 如有林窗存在, 幼苗迅速生长补充进现有种群, 如无林窗幼苗则很快死亡。这类树种, 种子寿命较短, 在萌发季节前的种子库中有活力种子, 萌发季节内出现幼苗, 但由于群落郁闭没有林窗, 幼苗当年存活率为零, 而在林窗处则有幼苗存活, 萌发季节后的种子库中没有活力种子, 也没有结构完整而无活力的种子, 如鞘柄菝葜。(3)种子成熟入土后, 如有林窗则迅速萌发更新到现有种群中去, 如无林窗则种子很快丧失活力。这类树种, 种子寿命较短, 在萌发季节前的种子库中有活力种子, 萌发季节内由于现存群落郁闭大而无林窗, 没有幼苗出现, 而林窗处有幼苗产生。在萌发季节后的种子库中无活力种子, 但有大量结构完整而无活力的种子。如樟叶槭、掌叶木、黄连木、黄棉木等。(4)种子成熟入土后, 如有林窗则迅速萌发更新补充到现有种群中去, 如无林窗则以种子的形式等待林窗出现。这类树种, 种子寿命较长, 在萌发季节前、萌发季节内及萌发季节后的种子库中均有活力种子, 很少或没有结构完整而无活力的种子, 在现有郁闭大的群落内萌发季节没有幼苗出现, 而在林窗处则有幼苗产生, 如圆果化香。
|
|

仔细分析种子库及其天然萌发可知, 茂兰喀斯特森林主要树种的繁殖与更新采取以上4种方式之一, 或兼而有之, 如羊蹄甲采取(1)、(4)方式, 多脉榆采取(1)、(3)方式, 圆叶乌桕采取(2)、(3)方式, 云贵鹅耳枥和朴树采取(2)、(4)方式。
4 讨论Michale Fenner(1985)从资源分配原则、繁殖能力、生活史与繁殖时间、种子的大小和数量等方面讨论过植物的繁殖方式, 但仅涉及繁殖, 而没有讨论其后的更新。朱守谦在讨论喀斯特森林群落的自我更新时, 认为喀斯特森林组成树种的繁殖更新方式有两种类型, 阴性树种以大量幼体处于等待状态; 阳性树种经常产生大量种子形成幼苗, 但在林下相继死亡而无较大幼树, 只有光照条件改善后才能形成幼树(朱守谦, 1993)。本研究认为喀斯特森林主要树种的繁殖更新方式有4种类型, 方式(1)和方式(2)相当于朱守谦讨论的两种方式。在这4种方式中, 阴性常绿树种趋向方式(1), 阳性落叶树种趋向方式(4)。树种的繁殖更新方式并不一定绝对附合某一方式, 有些树种可能几种方式兼而有之。
作者曾对贵州常态地貌上的常绿阔叶林进行过研究, 发现栲树群落内采取(1)、(2)、(3)、(4)方式的树种(包括兼行(1)、(2)、(3)、(4)方式者, 下同)所占比例分别为63.64 %、18.18 %、9.09 %和9.09 %。茂兰喀斯特森林群落内采取相应方式的树种所占比例分别为43.75 %、18.75 %、37.50 %和18.75 %。由此可知, 喀斯特森林与常态地貌上的常绿阔叶林相比, (1)方式下降, (3)方式和(4)方式增加。
森林树种繁殖更新方式的讨论, 有助于深入了解森林群落更新演替动态。以前往往从群落更新层中幼苗幼树状况来判断群落的发展趋势, 没有考虑到种子库中活力种子对群落发展的作用。实际上不能简单地认为群落更新层中没有幼体的种类就一定将被淘汰, 当老树死亡等原因形成林窗时, (2)、(3)、(4)方式的树种会迅速形成幼苗幼树补充进群落中去。
植物群落随着演替的进展, 群落郁闭度越来越大, (2)、(3)、(4)方式的更新越来越受到限制, 它们会逐渐减少。但喀斯特森林生境条件严酷, 土被不连续, 裸岩率高, 树木自然衰亡的年龄低, 经常有树木死亡形成林窗(周政贤, 1987)。本研究所设样地无林窗, 只是为了研究的需要而寻找的局部地段, 实际上茂兰大面积的喀斯特森林内, 林窗的存在是普遍现象。因此各种方式的树木能在茂兰喀斯特森林内长期存在, 从而使该类森林群落保持相对稳定。
植物繁殖更新方式的深入研究, 有助于生物多样性及珍贵树种的保护。由于有些植物要借助林窗才能很好更新, 林窗的经常产生有助于生物多样性的保持或增加, 有助于珍贵树种种群的扩大, 如二级保护植物掌叶木的繁殖更新采取(3)方式, 林窗的存在对其更新有利。现代生态学把处于两种或两种以上的物质体系、能量体系、结构体系、功能体系之间所形成的界面, 以及围绕这个界面向外延伸的过渡地带称为“Ecotone”地带。这个地带存在着一种边缘效应, 在自然条件下通常生物群落结构复杂, 出现多种生物共生的现象(于志熙, 1992)。林窗的出现就形成了这种过渡地带, 在这种地带内存在多种生态条件, 从而使各种繁殖方式的植物都能存在((1)方式在林内实现更新, (2)、(3)、(4)方式在林窗处实现更新)。因此在植被过密的地方, 人们对植被进行适当的开发利用形成一定的林窗, 能保护和增加生物多样性, 但林窗的大小及多少尚有待进一步研究。
刘济明. 1996. 梵净山栲树种群结实特征研究. 四川师范学院学报(自然科学版), 17(3): 20-23. |
刘济明, 雷基祥. 1997. 喜树种子的天然扩散. 贵州农学院学报, 16(2): 13-19. |
刘庆洪. 1988. 红松阔叶林中红松种子的分布及更新. 植物生态学与地植物学学报, 12(2): 134-142. |
于志熙. 1992. 城市生态学. 北京: 中国林业出版社, 90.
|
钟章成. 1988. 常绿阔叶林生态研究. 重庆: 西南师范大学出版社, 281-296.
|
周政贤. 1987. 茂兰喀斯特森林科学考察集. 贵阳: 贵州人民出版社, 1-224.
|
祝宁, 王义弘. 1992. 剌五加生殖生态研究(2)———种子扩散、种子库及更新. 东北林业大学学报, 20(5): 12-17. |
朱守谦主编.喀斯特森林生态研究(1).贵阳: 贵州科技出版社, 1993, 1~11
|
朱守谦主编.喀斯特森林生态研究(2).贵阳: 贵州科技出版社, 1997, 108~117
|
Harper J L. 1997. Population Biology of Plant. New York: Academic Press, 33-151.
|
Michale Fenner. 1985. Seed Ecology. London: Chapman and Hall, 57-116.
|
Silvertown J W. Introduction to Plant Population Ecology. London and New York, 1982, 22~37
|
Thompson K, Grime J P. 1979. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitat. Journal of Ecology, 67: 893-921. DOI:10.2307/2259220 |
Willson M F. Plant Reproductive Ecology. Willey, New York, 1983, 177~244
|