 2000, Vol. 36
2000, Vol. 36文章信息
- 田呈明, 康振生, 李振岐, 赵彦修, 张慧.
- Tian Chengming, Kang Zhensheng, Li Zhenqi, Zhao Yanxiu, Zhang Hui.
- 落叶松—杨栅锈菌遗传分化的RAPD分析
- ANALYSIS OF THE GENETIC SPECIALIZATION OF MELAMPSORA LARICI-POPULINA WITH RAPD MARKERS
- 林业科学, 2000, 36(5): 54-58.
- Scientia Silvae Sinicae, 2000, 36(5): 54-58.
-
文章历史
- 收稿日期:1999-09-13
-
作者相关文章


2. 山东师范大学 济南 250014
2. Shandong Normal University Jinan 250014
落叶松-杨栅锈菌(Melampsora larici-populina Kleb.)为害青杨派、黑杨派及其杂交种的多种杨树, 广泛分布于世界各杨树栽培区, 是杨树锈病中分布最广、危害最大的一种病害(袁嗣令, 1998; 周仲铭, 1985;Pino et al., 1997)。长期以来, 主要依赖化学药剂、选育和应用抗病品种来加以控制, 但随着病原菌新的生理小种的出现, 促使品种的抗病性降低或丧失。近年来, 已对不同生理小种与不同杨树无性系之间互作时的致病性差异进行了研究, 并从抗病性方面证明有7个毒性基因存在, 且用RAPD、AFLP技术标记了与抗性相关的基因位点(Lefever et al., 1998; Villar et al., 1996)但仍缺乏对病原菌群体分化的遗传分析, 对该菌生理小种的演化关系了解甚少。而研究病原菌种群的遗传分化可为杨树抗病育种和揭示病菌的毒性演化规律及病害防治提供理论基础。
传统的生理小种鉴定方法有费时费工、接种条件不易掌握以及鉴别品种的限制等缺陷, 难以满足生产需要。DNA随机扩增多态性(RAPD)可在分子水平上快速、灵敏地检测生物种群内的遗传分化, 国内外已在茄病镰刀菌(Fusarium solani f.sp.cucurbitae)、小麦条锈病(Puccinia striiformis f.sp.triici)、松干锈病(Cronartium spp.)等多种植物病原菌遗传多样性的研究上应用并获得成功。本文用RAPD技术对落叶松-杨栅锈菌不同地理来源的菌株进行了研究。
1 材料和方法 1.1 菌株来源供试菌株以夏孢子为材料。13个菌株分别采自陕西、青海两省的7个地区的卜氏杨、青杨和小叶杨(表 1)。采集带有夏孢子的新鲜病叶, 装入冰壶带回。采用李振岐等(1989)的方法, 将各菌株经单孢子堆分离并扩大繁殖后, 收集夏孢子并干燥15~20d后, 在真空安培管中置于低温下保存备用。
|
|
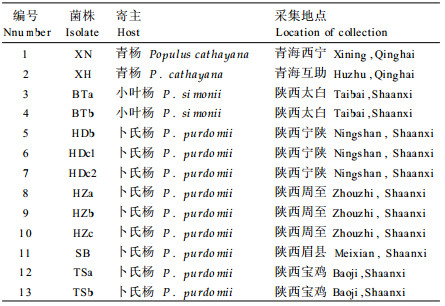
接种品种为美洲黑杨(P.deltoides)、欧美杨(P.× euramericana)、毛果杨(P.trichocarpa)、小叶杨(P.simonii)、青杨(P.cathayana)、卜氏杨(P.purdomi)、44 -114杨(P.deltoides × P.trichocarpa)等杨树品种的1年生扦插苗和2年生实生苗。
1.2.2 致病性测定1998~1999年的4~6月份分别将上述菌株经12 h水化处理后, 接种卜氏杨进行繁殖, 用繁殖的新鲜菌株涂抹法接种叶片(李振岐等, 1989), 每个品种接4盆(150片叶), 1盆为对照。接种时在叶片上喷上一层水膜, 用手指蘸取新鲜夏孢子悬浮液(5 ×106个夏孢子/mL), 均匀涂抹于叶背, 接种苗保湿24 h后, 在18℃~23℃培养, 每隔12 h定期观察并记载各处理的潜育期、反应型和单位面积(cm2)的夏孢子堆个数(重复统计3次, 取平均值), 寄主反应型分级标准参照曹支敏等(1998)制定的标准。
1.3 DNA提取DNA提取采用朱衡(1994)、单卫星(1995)的方法(并改进)。取50mg锈菌夏孢子, 液氮研磨至粉状, 加到装有500 μL预冷提取液(100 mmol/L Tris -HCl, pH 9.0;50 mmol/L EDTA, pH8.5)的1.5 mL离心管中, 振荡混匀后, 加入50 μL 20% SDS, 混匀后于65℃水浴50 min(每10 min混匀1次)。加入等体积氯化苄, 振荡混匀后, 65℃保温30min。再给混合液中加入1/10体积预冷的3 mol/L乙酸钠溶液(pH 5.2), 冰浴1 h, 15000 g下离心10 min, 取上清液并移入另一1.5 mL离心管中, 加入等体积的酚:氯仿(1:1), 10000 g离心10 min, 取上清液并加入2 μL RNase(10ug/μL), 37℃保温30 min, 加氯仿抽提1遍, 取上清液加入等体积的异丙醇混合, -20℃静置30 min沉淀DNA, 然后10000 g下离心10 min, 沉淀用70%的乙醇和无水乙醇依次漂洗后, 风干, 加50 μL TE(10 mmol/Tris -HCl, 0.1 mmol/L EDTA, pH8.0)溶解DNA, …20℃保存备用。
1.4 PCR扩增DNA扩增在Perkin-Elmer DNA Therm al Cycler 480型PCR仪上进行。扩增随机引物购自美国Operon公司。扩增反应为25 μL体系:200 μmol/L dN TP(N=A, C, G, T), 0.15 μmol/L随机引物, 2.5μL 10X扩增缓冲液(100mmol/L Tris -HCl, pH8.3;500mmol/L KCl, 20 mmol/L MgCl2; 0.001%明胶), 1.0U Taq酶(本室自制), 10ng模板DNA, 超重水补至25 μL, 并覆盖适量矿物油。反应程序:94℃1 min, 35℃ 1 min, 72℃ 2 min, 反应5个循环后, 然后94℃ 20 s, 35℃ 40 s, 72℃ 1 min, 反应40个循环, 最后72℃延伸10 min。扩增产物在含有溴化乙锭(0.5 μg/mL)的1.5%琼脂糖胶中电泳检测分析。
1.5 数据分析对电泳谱带进行记录, 同一位置有带的记为1, 无带的记为0, 统计各菌株的扩增条带数, 并根据公式S=2 Nab/(na +nb), 计算各菌株间的相似系数。用NTSYS(Version 1.70, 1992)软件中的非加权平均连锁法(UPGMA)进行聚类分析, 并构建系统进化图。
将供试菌株在鉴别寄主上的反应型、潜育期及产孢量统计后, 用非加权平均连锁法(UPGMA)进行聚类分析, 并构建系统进化图, 与RAPD结果进行比较分析。
2 结果与分析 2.1 致病性测定结果用13个菌株对7个杨树鉴别品种接种后, 依据分级标准(曹支敏等, 1998)记载反应型如下:
从各寄主对不同菌株的反应型(表 2)可以看出, 美洲黑杨(P.deltoides)对13个菌株均表现为近免疫; 毛果杨(P.trichocarpa)、44-117杨(P.deltoides ×P.trichocarpa)均表现为高感; 小叶杨(P.simonii)、卜氏杨(P.purdomi)表现为中感~高感反应; 不同菌种在欧美杨(Populus × euramericana)和青杨(P.cathayana)上的反应型差异较大, 从高感到高抗; 在青杨上, 13个菌株中的1~4和11~13为高抗, 5~7为中感, 8~10为高感; 在欧美杨上也可分为1~4(中感), 5~10(高感), 11~13(高抗)等3个类群。
|
|
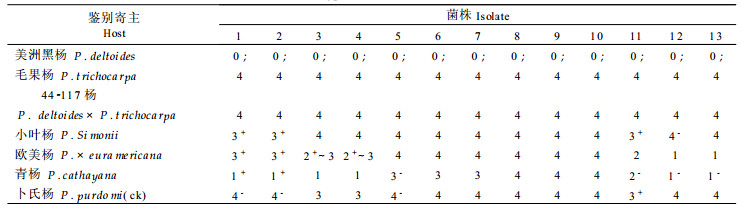
接种后定期观察记载潜育期及夏孢子堆数量(表 3)发现, 亲和性(感病性)反应的潜育期较短(4~6d), 产孢量大(15~103个/cm2); 不亲和反应(抗病性)的潜育期较长(7~9 d), 产孢量小(1~15个/cm2); 近免疫品种的潜育期为13~16 d, 主要表现为轻度褪绿斑, 不产孢, 也不产生枯斑。
|
|
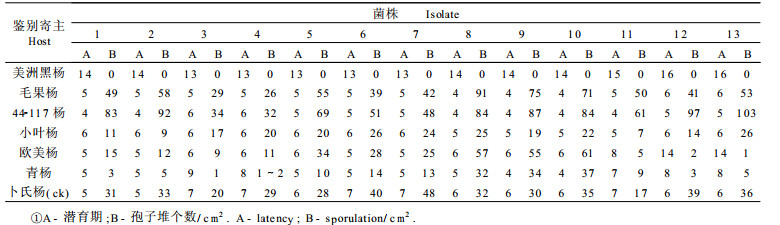
根据反应型、产孢量及潜育期对13个菌株的聚类结果显示(图 1), 所有菌株被分为3个类群:即来自西宁的菌株XN、XH与天台山的菌株TSa、TSb为类群Ⅰ; 火地塘的菌株HDc1、HDc2、HDb与厚畛子的菌株HZa、HZb、HZc为类群Ⅱ; 宝太路(BAa~b)、上白云(SB)等地的菌株为类群Ⅲ。

|
图 1 M.larici-populina13个菌株致病性聚类分析树状图 Fig. 1 UPGM A dendrogram based on virulence data from 13 isolates of M.larici-populina |
用80个引物(KI Ts:C01~20, G01~20, H01~20, U01~20), 对供试各菌株的DNA混合后(DNA pool)进行扩增, 根据筛选结果选出13个重复性好、特异性强且谱带清楚的随机引物。用13个随机引物对13个菌株进行RAPD扩增, 产生81条DNA带, 其中69条呈现多态性, 多态性带数占总带数的56林业科学36卷85.2%(表 4, 图 2)。扩增带数为3~11条。扩增片段在0.5~2.5 kb之间。
|
|


|
图 2 Melampsora larici-populina不同菌株的RAP D扩增图谱(引物OPH08和O PH12) Fig. 2 RAPD bands of 13 isolates of M.larici-populina treated with primer OPH08 and OPH12 |
13个引物对来自不同地区及不同杨树品种上的落叶松-杨栅锈菌的PCR扩增谱带的聚类结果显示, 所有供试菌株的相似系数在0.608~1.000之间, 各菌株之间的差异在0~33.1%之间, 说明不同菌株之间存在着较丰富的多态性。聚类结果(图 3)显示, 13个菌株可明显分为4个类群:第1类群包括H Dc1、HDc2, H Dc1和HDc2为卜氏杨的一个叶片上的两个单孢子堆分离物; 第2类群为HDb; 第3类群为来自青海的2个菌株XN、XH和秦岭北坡的6个菌株BTa、BTb、HZa、SB、TSa、TSb, 在相似性系数0.791时被聚为1类; 第4类群包括HZb和HZc两个分离物。

|
图 3 M.larici-populina 13个菌株的RAPD聚类UPGMA树状图 Fig. 3 UPGMA dendrogram based on RAPD data from 13 isolates of M.larici-populina |
RAPD分析发现一些来源于同一地点的不同分离系之间, 也存在较高的遗传距离。如HDc1、HDc2与HDb, HZa与HZb、HZc以及这两个地点的菌株之间都存在较高的遗传距离, 但同一地点各菌株以及火地塘(HD)与厚畛子(HZ)两地菌株的致病性极为相似。按致病性分析XN、XH与TSa、TSb, BTa~b与SB的关系较近, 而RAPD分析结果显示XN、XH、BTa、BTb、TSa、TSb、SB、HZa等菌株具有较高的遗传相似性, 但各菌株之间以DNA标记的遗传距离也存在差异。
3 讨论落叶松-杨栅锈菌的生理分化现象在1949年由Van Vloten报道了发生于荷兰的3个生理小种和1个变种, 但直到70年代末和80年代初, 国外学者才对该菌的生理分化、致病性等进行了较为系统的研究, 并命名了E1、E2、E3、E4等4个生理小种。上述研究主要用鉴别寄主来划分生理小种, 而目前依据生物化学、分子生物学性状区分生理小种, 已成为植物锈菌小种研究的重要手段, 但在树木锈菌研究方面尚不多见。曹支敏等(1998)依据鉴别寄主及可溶性蛋白将采自陕西秦岭的落叶松-杨栅锈菌(M.larici-populina)划分为3个生理小种:MLP1、MLP2和MLP3。
本研究应用RAPD技术对松杨栅锈菌(Melampsora larici-populina Kleb.)生理分化进行了研究, 并和致病性结果进行了比较, 发现该菌存在明显的生理分化现象。供试菌株在鉴别寄主上的致病性聚类分析树状图与其各遗传距离划分的RAPD指纹组之间的相关性分析表明, 病原菌的致病性与病菌本身的DNA遗传分化之间有一定差异, RAPD标记的DNA多态性与菌株的毒性多态性相关性不明显, 这可能与锈菌的高度寄生性有关, 与Cheng(1993)、单卫星(1995)、朴春根(1996)等对小麦锈病的研究结果相似。
来自不同地理环境和不同寄主的菌株的遗传分化没有明显差异, 如来自青海的青杨上的2个菌株(XN、XH)和陕西秦岭北坡卜氏杨、小叶杨的6个菌株(BTa、BTb、HZa、SB、TSa、TSb)的相似性为79.1%, 遗传距离较近, 显示出菌株的DNA多态性与寄主种类和地理环境之间没有相关性。按致病性特征HZa与采自同一地区的同一树种的HZb、HZc的亲缘关系较近, 但RAPD分析发现HZa与HZb~ c遗传距离较远。但来自秦岭南坡(火地塘)和北坡(天台山、上白云)卜氏杨上的菌株显示较大的差异, 其原因有待进一步分析。
RAPD分析是以基因组DNA随机位点的差异为基础, 而致病性则与致病基因编码的基因组相联系(包括多个致病基因及同一致病基因的不同位点), 并受寄主基因组的强烈选择作用。本研究仅就青海和陕西地区的M.larici-populina菌株的遗传分化进行了分析, 然而, 由于供试菌株数量的限制, 难以对DNA多态性与致病性的相关性作进一步分析, 对于病原菌群体的变异规律及其与毒性的关系需进一步研究。
程东升, 梁惠燕, 薛煜, 等. 1998. 中国三种松干锈菌在随机扩增DNA多态性水平上的遗传分化. 林业科学, 34(5): 53-59. DOI:10.3321/j.issn:1001-7488.1998.05.007 |
曹支敏, 李振岐, 胡景江. 1998. 落叶松-杨栅锈菌生理分化研究. 西北林学院学报, 13(1): 53-57. |
韩正敏, 尹佟明, 黄敏仁, 等. 1998. 用RAPD研究我国杨生褐盘二孢菌的群体分化. 植物病理学报, 28(4): 347-352. DOI:10.3321/j.issn:0412-0914.1998.04.013 |
李振岐, 商鸿生. 1989. 小麦锈病及其防治. 上海: 上海科学技术出版社, 211-220.
|
单卫星, 陈受宜, 吴立人, 等. 1995. 中国小麦条秀菌流行小种的RAPD分析. 中国农业学报, 25(5): 1-7. |
朴春根, 唐文华, 曾士迈, 等. 1996. RAPD技术和局类分析在小麦条锈病菌生理小种研究中的应用. 植物病理学报, 26(3): 205-210. DOI:10.3321/j.issn:0412-0914.1996.03.002 |
田呈明, 李振岐, 康振生. 1999. 青杨叶锈病(Melampsora larici-populina Kleb.)研究进展. 西北林学院学报, 14(3): 81-88. DOI:10.3969/j.issn.1001-7461.1999.03.015 |
袁嗣令. 1998. 中国乔灌木病害. 北京: 中国林业出版社, 110-112.
|
周仲铭, 袁毅. 1985. 杨树叶锈病的研究概况. 北京林学院学报, (4): 84-102. |
朱衡, 瞿峰, 朱立煌. 1994. 利用氯化苄提取适于分子生物学分析的真菌DNA. 真菌学报, 13(1): 34-40. |
Pino J, Frey P. 1997. Structue of Melampsora larici-populina populations on wild and cultivated poplar. Eur.J.Plant Pathol., 103(2): 159-173. |
Lefever F, Goue Mourier M C, Faivre Rampant P, et al. 1998. A single gene cluster controls incompatibility and partial resistance to various Melam psora larici-pop ulina races in hybrid poplars. Phytopathology, 88(2): 156-163. DOI:10.1094/PHYTO.1998.88.2.156 |
Villar M, Lefevre F, Bradshaw HD Jr, et al. 1996. Molecular genetics of rust resistance in poplar (Melampsora larici-populina Kleb./ Populus sp.)by bulked segregant analysis in a 2×2 factorial mating design. Genetics, 143(1): 531-536. |