 2000, Vol. 36
2000, Vol. 36文章信息
- 张小全, 徐德应.
- Zhang Xiaoquan, Xu Deying.
- 杉木中龄林不同部位和叶龄针叶光合特性的日变化和季节变化
- SEASONAL CHANGES AND DAILY COURSES OF PHOTOSYNTHETIC CHARACTERISTICS OF 18-YEAR-OLD CHINESE FIR SHOOTS IN RELATION TO SHOOT AGES AND POSITIONS WITHIN TREE CROWN
- 林业科学, 2000, 36(3): 19-26.
- Scientia Silvae Sinicae, 2000, 36(3): 19-26.
-
文章历史
- 收稿日期:1999-01-11
-
作者相关文章


树木光合作用对CO2的同化是森林生态系统能量流动和物质循环的基础, 是决定森林生产力的最重要的生理过程; 同时光合作用及其生产力的形成又受外界各种环境因素和树木自身光合特性的影响。不少研究人员对杉木针叶光合作用进行过研究(俞新妥, 1989; 郭继善, 1992; 吴立勋等, 1992; 陈洪, 1995; 范少辉等, 1996), 并涉及到对不同叶位(郭继善, 1992; 吴立勋等, 1992)和叶龄针叶(郭继善, 1992; 陈洪, 1995)光合特性的比较研究。但多数研究测定是在随机或者无法控制的环境条件下进行, 测定结果差异较大, 不同研究结果之间的可比性差。本文系统研究测定了不同生长季节光合最适温度、适宜湿度和正常大气CO2浓度条件下, 杉木中龄林不同部位和叶龄针叶的光合特性及不同天气下的日变化特征, 以期为冠层光合生产力模拟提供必要的信息。
1 试验地概况与研究方法 1.1 试验地概况研究林分位于江西省分宜市亚热带林业实验中心年珠实验林场, 地理位置114°33′47″E, 27°34′41″N, 是杉木分布的中带中部产区。地带性植被为常绿阔叶混交林。气候属亚热带湿润气候区。年均气温16.8℃, 极端最高温39.9℃, 极端最低温-8.3℃, 年积温5355℃; 年降水量1590 mm, 年蒸发量1504 mm; 年均日照1657 h; 太阳辐射年总量约487 KJ/cm2; 年均相对湿度80%;无霜期270 d。试验样地海拔240~270 m, 坡向南偏东20°, 坡度20°。成土母质为砂页岩, 土壤属黄棕壤, 土层厚度70~100cm。林分为杉木人工纯林, 林龄18 a, 地位指数16, 株行距2.0 m×1.5 m, 平均胸径13.8 cm, 平均树高12.6 m, 平均冠幅2.89 m, 枝下高7.52 m。
1.2 野外测定方法 1.2.1 光合特性测定在样地内根据林分平均胸径、平均树高、平均冠幅和冠长, 选取1株标准木, 将树冠等分为上、中、下三层, 在每层向南伸展的侧枝上选择当年生、1年生和2年生(包括2年生以上, 以下简称2年生)代表性样枝各一枝, 分别于1998年生长盛期(5月下旬)、中期(7月下旬)和后期(10月中旬), 在找出不同季节、不同叶龄针叶最适温、湿度范围的基础上(已另文叙述), 在正常大气CO2浓度、最适湿度范围、相对湿度70%±5%左右条件下, 测定不同光合有效辐射通量密度(PPFD)下针叶的净光合速率(Pn)以及暗呼吸(Rd)。测定是在叶室中以活体小枝为单位进行的。所用仪器为LiCor-6200光合测定系统, 但对其4 L叶室进行了改装, 通过水循环和温控器来稳定和调节叶室内气温。PPFD通过遮盖不同厚度白色沙布的方法来调节, 当太阳PPFD不足时, 用400W镝灯作为辅助光源。在1998年5~10月全部测定期间雨量充沛, 未发生土壤水分亏缺, 因此可以认为土壤水分供应充足。
1.2.2 日变化测定用上述方法选择样枝, 在不加任何人为环境控制的自然状态下测定不同部位和叶龄针叶Pn的日变化。日出后30 min开始, 每2 h测定1次。测定时叶室水平放置。
1.2.3 光合叶面积测定杉木针叶为披针形片状体, 且单面有气孔, 故小枝针叶面积可采用复印称重法测定, 即将测定枝上的针叶全部剪下, 用透明胶带粘贴于复印纸上复印, 将复印出的针叶剪下称重, 根据单位复印纸的重量计算测定枝上的叶面积。
1.3 数据统计分析不同季节、不同部位和不同叶龄针叶Pn与PPFD之间的关系, 用最小二乘法采用下述经验方程进行拟合(Bassman等, 1991):
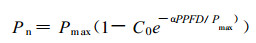
|
(1) |
式中Pmax代表最大净光合速率, α为弱光下光量子利用效率(即表观量子效率), C0为一度量弱光下净光合速率趋近于零的指标。通过适合性检验, 若方程拟合效果良好, 则可用下式计算光补偿点(LCP):

|
(2) |
假定Pn达到Pmax的99%时的PPFD为光饱和点(LSP), 则:
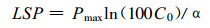
|
(3) |
通过对不同生长季节不同部位和叶龄27个杉木针叶样枝的测定和拟合, 结果列于表 1。由于篇幅所限, 文中未列出测定的27个样枝的散点图和拟合曲线, 不同季节的比较仅列出中部针叶(图 1), 不同叶龄和部位针叶的比较仅列出生长中期的情况(图 2, 图 3)。

|
图 1 不同季节光响应曲线比较(树冠中部) Fig. 1 Comparisons of PPFD responses of net photosynthetic rate (Pn) for 3 growing seasons at mid-crown ……盛期 Fast-growing season; ———中期 Mid season; -·-·后期 Late season. |

|
图 2 不同叶龄针叶光响应曲线比较(生长中期) Fig. 2 Comparisons of PPFD responses curves of Pn for different age of shoots in mid season ……当年生 Current, ———1年生 1-year-old, -·-·2年生 2-year-old. |

|
图 3 不同部位针叶光响应曲线比较(生长中期) Fig. 3 Comparisons of PPFD responses of Pn for shoots at different positions within crown in mid season ……上部 Upper crown, ———中部 Middle crown, -·-·下部 Lower crown. |
在最适温度和较适宜湿度条件下得出的光饱和最大净光合速率(Pmax)是衡量针叶光合能力的重要指标。表 1和图 1表明不同季节针叶Pmax不同。生长盛期(5月下旬)杉木处于生长旺期, 生理活性强, 因而净光合速率较高。从生长盛期至中期到后期Pmax呈降低趋势。但当年生针叶例外, 其中期的Pmax值明显高于盛期, 这与生长盛期当年生叶尚未发育成熟、光合机构未完善和呼吸作用强有关。
|
|

不同叶龄杉木针叶Pmax不同(表 1, 图 2)。除了在生长盛期由于当年生针叶尚未发育成熟, 其Pmax与1年生针叶相近, 其它均表现出当年生大于1年生, 而1年生明显大于2年生。
在树冠不同部位, 针叶Pmax差异显著(表 1, 图 3)。无论在生长盛期、中期、后期, 还是当年生、1年生、2年生针叶, 从树冠上部到下部, Pmax呈下降的趋势。相对而言, 上部与中部之间的差别较小, 而上、中部与下部之间Pmax的差别则明显得多。
2.2 光补偿点(LCP)、光饱和点(LSP)和表观量子效率(α)LCP、LSP和α是指示植物光响应特征的重要指标。LCP和LSP均可通过前述光响应方程计算。从实际拟合效果看, 拟合的光响应曲线比实测值较早稳定下来, 这是这种类型曲线所决定的, 因此计算出的LSP比实际要低。所以我们用实测点的走向来估计LSP的值。LCP则采用上述计算值(表 1)。
表 1和图 1~3表明, 不同季节和叶龄针叶α、LCP和LSP不同。从生长盛期经中期到后期, LSP有降低的趋势, α则呈增加趋势, 表明针叶对强光的利用能力下降而对弱光的利用能力增强。其中生长盛期与中、后期差别较大, 而中期与后期差异不明显。LCP除生长盛期当年生针叶明显高于生长中、后期以外, 其它差异不明显。就不同叶龄针叶而言, 当年生针叶具有较高的LCP和LSP, 其次为1年生针叶, 2年生针叶LCP和LSP最低; 而表观量子效率则相反。也就是说, 随着针叶年龄的增长, 针叶LSP和LCP降低, α提高, 表明针叶对弱光的利用能力增强, 对强光的利用能力减弱。
不同部位针叶LSP、LCP和α也存在明显差异, 在同一生长季节相同叶龄的针叶, 上部针叶LCP和大于中部下部最低而的变化正好相反但上部与中部的差异相对较小而上中部与下部的差异较大。说明树冠下部针叶具有较强的弱光利用能力, 而中上部针叶具有较强的强光利用能力。
2.3 暗呼吸速率(Rd)在光合最适温度范围、正常大气CO2浓度和70%±5%相对湿度下测得的不同季节和部位杉木针叶暗呼吸速率表明(表 1), 不同针叶年龄, 暗呼吸有明显差别。尤其是当年生针叶, 其呼吸速率显著高于1年生和2年生老叶。就不同部位而言, 上部针叶呼吸作用最强, 中部次之, 下部最弱。不同季节呼吸速率不同, 表 1中期的呼吸速率大于盛期和后期, 但是中期测定时的温度较高(30±1℃), 而盛期和后期较低(22±1℃或25±1℃), 所以如果利用测定和拟合的暗呼吸对温度的响应曲线(已另文叙述)换算到相同温度条件下, 则表现出盛期大于中期, 中期大于后期的趋势。
2.4 光合作用日变化特征植物光合作用受各种生态、生理和生化因素的影响和制约, 而这些因素无时无刻不在发生变化, 使光合作用呈现出复杂的季节变化和日变化。对生长中期两个典型天气(晴天和阴天)条件下不同部位和叶龄针叶净光合速率的连续测定表明(图 4), 杉木中龄林针叶净光合速率的日变化在阴天为单峰型, 而晴天为双峰型。在阴天条件下, 净光合速率的日变化节律似乎与辐射的变化十分吻合, 即早晨随辐射的增加而增大, 于10:00~11:00时达最大值, 以后随辐射的减弱, 净光合速率也下降, 在15:00以后, 辐射有所增强, 净光合速率也有所增加(图 4-A~B)。在晴天条件下, 净光合速率在一天中有两个峰值, 一次出现在上午9:00左右, 另一次在下午15:00时左右, 尽管中午太阳辐射很强, 但净光合速率明显较低(图 4-D~E)。

|
图 4 不同部位和叶龄针叶净光合速率(Pn)及相关因子的日变化(A~C为1997年7月20日, 阴; D~F为1997年7月14日, 晴) Fig. 4 Daily courses of net photosynthetic rate (Pn) and related environmental factors for shoots with 3 age classes at 3 positions within crown (A~C:July 20, 1997, Cloudy; D~F:July 14, 1997, Clear)
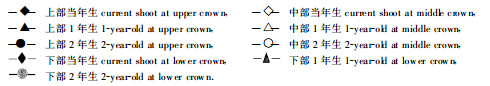
|
就不同部位和叶龄针叶而言, 当年生新叶和中上部针叶光合作用具有明显的日变化特征, 而中下部和老叶, 特别是2年生老叶, 日变化幅度较小, 甚至在晴天条件下只有一个峰值(图 4-A, D)。然而, 尽管不同部位和叶龄针叶日变化有差异, 但由于受本身生理活性和辐射条件的作用, 净光合速率的绝对值仍是当年生大于年生年生大于年生上部大于中部中下大于下部与前述变化趋势相似
3 讨论 3.1 最大净光合速率Pmax同生长一样, 杉木的光合生理活性也具有明显的季节变化, 且除当年生针叶略有不同外, 与杉木生长节律基本一致。范少辉等(1996)测定表明, 净光合速率在6月份最高, 9月次之, 4月和11月最低, 与本研究中老叶的季节变化基本相似。
不同部位针叶Pmax之间的差异, 与郭继善(1992)对12年生杉木针叶的测定相似, 且与肖文发(1994)对该样地年生杉木同一枝条上不同部位针叶的测定结果相似即枝前部大于枝中部大于枝后部。但Pmax低于肖文发1)的测定值, 可能因为本研究为生长明显处于衰退期的18年生林分, 而13年生杉木正处于生长旺盛期, 生理活性较强。范少辉等(1996)测定表明, 10年生杉木针叶的净光合速率明显大于5、15和19年生。吴立勋和徐世凤(1992)也发现, 12年生杉木阳位叶净光合速率明显大于阴位叶。不同部位针叶光合能力的差异与针叶长期适应所处的光环境有关(苘辉民, 1994)。杉木中龄林具有较大的冠长和冠径, 不同部位针叶所处的光环境不同。通常上部针叶光照条件优于中部, 而下部针叶长期处于被遮荫状态, 一天中很少时间能获得较高的辐射。
1) 肖文发.杉木人工林冠层光合生产力的模拟研究.博士论文, 1994。
除生长盛期当年生针叶外, 不同季节和部位针叶光合能力均表现出当年生大于1年生, 1年生大于2年生的趋势。但一些测定表明, 2年生针叶净光合速率最大(郭继善, 1992; 陈洪, 1995), 值得商榷。一般而言, 树叶在生长、成熟直至衰老过程中, 其光合能力是不断变化的, 当木本植物的新枝条开始伸展时, 可以测出整个白天在光下有CO2的净释放; 随着叶子的展开, CO2固定能力很快增强。新发育成熟的叶片光合能力达到高峰, 并在几天或几个星期后, 光合能力便降低了, 随着植株变老而稳定地下降(苘辉民, 1994; 龚垒, 1989; Larcher, 1980)。本文和其它一些测定(吴立勋等, 1992)均证明了这一点。虽然本文未能在整个生长季节对当年生叶光合能力的变化进行连续的测定, 但在针叶发育期, 光合能力较低, 在发育成熟后达到最大, 这是不容置疑的。叶片光合能力之依赖于发育时期, 通常与叶绿素含量、酶活性和呼吸强度有关(龚垒, 1989; Larcher, 1980)。处于旺盛生长期的叶子, 发育不完全, 叶绿素含量低, 与光合作用有关的酶的活性低, 而呼吸作用旺盛, 因而光合作用受到限制。随着叶片和气孔的发育成熟, 叶片对CO2的扩散阻力下降, 叶绿素和蛋白质的合成活跃, 光合磷酸化速率和RUBP羧化酶活性增加而线粒体呼吸下降, 因而光合能力增强。随着叶片衰老, 上述几方面因素又朝着不利于光合作用的方向变化(龚垒, 1989)。
3.2 光饱和点LSP、光补偿点LCP和表观量子利用效率α在不同生长季节, 不同部位和叶龄针叶LSP、LCP和α均存在较大差异。不同季节的差异与其本身的生理活性如叶绿素含量、酶活性、气孔反应等有关, 不同部位之间的差异则与其长期所处的光环境有关。正如Larcher (1980)所述, LCP和LSP的值反映着植物在自然生境中所处的光照条件。而不同叶龄针叶之间的差异除与生理活性有关外, 还可能与针叶适应所处的光环境有关, 因为当年生一般处于树冠外层或枝顶端, 能受到充分的光照, 而老叶特别是2年生针叶常处于树冠里层, 光环境差。
与其它有关杉木光合特性研究结果相比较, 发现LSP低于肖文发的13年生和范少辉(1996)的19年生杉木林, 与前者的差异可能与林分年龄有关, 与后者的差异可能源于测定方法(离体测定)和林分地域(福建)的差异。但吴立勋等(1992)测定的12年生林分中上部当年生叶LSP (471~1413μmol m-2s-1)低于我们的测定, 可能与其测定方法有关(离体测定)。LCP与肖文发的结果相似, 但大大低于陈洪(1995)、范少辉(1996)及吴立勋等(1992)的测定值。本研究与其它有关杉木的测定差异除了地域、林龄、测定仪器和方法外, 测定时的微环境因素也可能是重要原因。不同地域的立地条件和气候因素不同, 杉木的生理活性可能存在较大差异。温度、CO2浓度、水肥条件和植物生长状况等都对LSP和LCP产生影响(苘辉民, 1994)。离体测定时尽管将枝条浸入水中, 但仍会使针叶水势降低, 产生水分胁迫从而使LCP提高和LSP降低。测定时的微环境如大气湿度和温度也会对测定结果产生影响, 较低的大气湿度和较高的温度都有可能提高LCP和降低LSP (龚垒, 1989; 郭连生等, 1994; 胡新生等, 1997)。
3.3 暗呼吸速率Rd在不同季节不同部位和叶龄针叶Rd存在明显差异。5月下旬是杉木生长高峰, 也是当年生针叶快速展叶期。由于幼嫩组织呼吸速率大于老组织, 正在生长的组织的呼吸速率大于成熟组织(Larcher, 1980; Bidwell 1979), 使盛期当年生针叶具有明显较高的Rd。此外, 不同叶龄针叶Rd的差异还可能与其本身所处的光环境有关, 从冠层上部到下部叶片的暗Rd随叶片接受辐射的减少而降低(Hagihara等, 1991)。老叶尤其是2年生及2年生以上针叶一般处于树冠的里层和下层, 受光条件较差, 而当年生叶则常处于树冠顶层或外围, 光环境优越。
3.4 光合作用的日变化杉木针叶日变化具有双峰型和单峰型两种形式, 与天气条件有关。从其它有关杉木光合作用日变化的测定结果看, 有双峰型(陈洪, 1995), 也有单峰型。但双峰型中, 陈洪(1995)测定的2个峰值分别出现在10:00~12:00时和16:00~17:00时, 均比我们的测定晚约2小时。肖文发的测定与我们的结果相近, 两个峰值分别在9:00~11:00时和13:00~15:00时, 单峰型的峰值在9:00~11:00时, 但其单峰型发生在秋季, 春、夏两季为双峰型。这些测定结果的差异, 可能与林分年龄、生长季节、天气条件、地形、立地条件等诸多因素有关。因此, 对于杉木针叶而言, 很难断定其日变化一定是单峰型、双峰型以及其峰值出现的早晚。目前的研究只能表明, 杉木针叶光合作用的日变化在某些条件下呈单峰型, 在某些条件下呈双峰型。
从本次测定情况看, 在阴天条件下(1997年7月20日), 测定时段最低相对湿度也在75%以上, 最高饱和水汽压差(VPD)为10 mb左右, 气温(25~31℃)基本在最适温度范围, CO2浓度340~380μl.l-1 (图 4-C)。根据我们的测定, 饱和水汽压差在8~10 mb以下, 或大气相对湿度在70%以上, 净光合速率变化不明显。可以推断, 该阴天的光合日变化主要与辐射有关, 同时受温度、湿度和CO2浓度等因子的微弱变化影响。在1997年7月14日的晴天条件下, 测定时段内大气相对湿度最高达95% (早晨), 最低58% (中午), 气温25~37℃, 气温和相对湿度变化引起的VPD变化在1~25 mb之间(图 4-F), 高温、低湿和高VPD会从气孔和非气孔方面限制光合作用。特别是上午10:00时至下午15:00时, 相对湿度在70%以下, VPD在10 mb以上, 气温在33℃以上(图 4-F), 导致气孔关闭和光合活性下降, 这可能是导致晴天中午光合速率下降的主要原因。此外在晴天长时间高光强下, 也可能发生光抑制, 从而导致光合速率的降低。
但是, 目前的研究还不能断定晴天中午光合速率的下降就是植物的“午睡”现象。因为“午睡”现象的发生与否及其原因, 不仅涉及光照、土壤水分条件、空气温度、大气湿度等生态因子, 还涉及气孔导度、叶肉阻力、叶片水势、呼吸作用、生理节奏等生理因素, 还可能与光合产物积累、RuBP羧化酶活性、光化学系统效率和脱落酸(ABA)含量等生化因子有关。目前对光合“午睡”的机理尚无定论(许大全, 1990)。
陈洪. 1995. 杉木幼林若干光合特性的研究. 福建林业科技, 22(3): 69-72. |
范少辉, 沈国舫, 等. 1996. 不同立地条件下不同林龄杉木人工林生理特性的研究, Ⅱ.光合特性. 林业科学研究, 9(mem.): 60-65. |
郭继善. 1992. 利用14CO2研究杉木的光合特性. 南京林业大学学报, 16(4): 86-89. |
郭连生, 田有亮. 1994. 4种针叶幼树的光合生理特性与大气湿度关系的研究. 生态学报, 14(2): 136-141. DOI:10.3321/j.issn:1000-0933.1994.02.001 |
苘辉民. 1994. 植物生理学. 北京: 北京农业大学出版社.
|
胡新生, 刘建伟, 王世绩. 1997. 4个树种无性系在不同温度和相对湿度条件下净光合速率的比较研究. 林业科学, 33(2): 107-116. DOI:10.3321/j.issn:1001-7488.1997.02.002 |
龚垒. 1989. 树木的光合作用与物质生产. 北京: 北京科学技术出版社.
|
吴立勋, 徐世凤. 1992. 杉木、檫树光合特性研究. 湖南林业科技, (2): 5-10. |
许大全. 1990. 光合作用"午睡"现象的生态. 生理与生化.植物生理学通讯, (6): 5-10. |
俞新妥. 1989. 不同种源杉木光合性状的比较研究. 福建林学院学报, 9(3): 223-237. |
Bassman J B and Zw ier J C. 1991. Gas exchange characteristics of Populus trichocarpa, Populus deltoides and Populus trichocarpa ×Pdeltoides clone. T ree Physiology, 8: 145-149. |
Bidwell R G S.Plant Physiology.M acM illan Publishing Co., Inc., New York, 1979
|
Hagihara A and Hozumi K.Respiration.In " Physiology of Trees" (A.S.Raghavendra, ed.), 87 -110, John W iley & Sons, INC.New York, 1991
|
Larcher W.植物生理生态学(李博等译).北京: 科学出版社, 1980
|