 2000, Vol. 36
2000, Vol. 36文章信息
- 裴保华, 袁玉欣, 贾玉彬, 王文全, Josef Eitzinger.
- Pei Baohua, Yuan Yuxin, Jia Yubin, Wang Wenquan, Josef Eitzinger.
- 杨农间作光能利用的研究
- A STUDY ON LIGHT UTILIZATION OF POPLAR-CROP INTERCROPPING SYSTEM
- 林业科学, 2000, 36(3): 13-18.
- Scientia Silvae Sinicae, 2000, 36(3): 13-18.
-
文章历史
- 收稿日期:1998-10-05
-
作者相关文章


2. Universität fǜr Bodencultur Wien Insititut fǜr Meteorologie und Phisik AUST RIA
2. Universität fǜr Bodencu ltur Wien Insititut fǜr Meteorologie und Phisik AUS TRIA
近年来我国利用世界银行贷款在黄淮海平原农区营造了大面积以林粮间作为主要模式的速生丰产林, 据统计仅河北省在第1、2期项目中已造林约7×104hm2这些林分多分布在沙荒地风沙危害的次耕地, 少数分布在重粘质土或轻盐渍土上。黄淮海平原长期受旱、涝、风沙、盐碱等不利自然因素危害, 农业产量低而不稳。实行农林复合经营, 是控制风沙, 改善环境, 充分利用自然资源的有效措施。
杨农间作, 杨树处于立体种植的上层, 它对农作物的遮荫, 是农作物减产的重要原因之一。研究间作田作物冠层表面光强变化, 及其与农作物光能利用率和产量的关系, 是调节杨农间作结构, 延长间作年限, 减轻负效应, 充分发挥立体种植优越性的重要依据。为此, 研究了密植幼林杨农间作田的光能分布, 作物和林木的日累积光合量和光能利用率; 比较分析了间作与对照的总光能利用率和农作物产量。
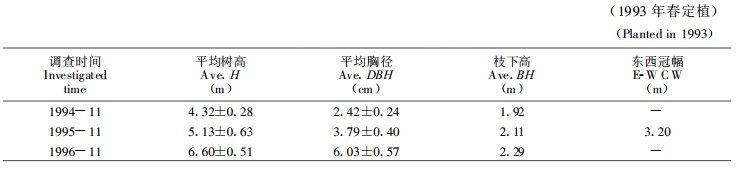
1 研究方法 1.1 杨农间作田林木生长情况试验地位于保定市郊区, 供试树种为白杨派双杂交优良无性系741杨{[ (Populus alba×P.Davidiana) +P.simonii]×P.tomentosai}, 林龄3~4年, 南北行向, 株行距0.5 m×5 m。这种密度只适用于杨树幼龄期的林粮间作。林木生长状况见表 1。
|
|
1995年6月20日用LCA-3光合测定系统测定间作田内外的光强(PAR)和光合速率; 7月20日、8月10日和1996年4月14日、5月14日和6月2日, 用ST-80C数字式棒状照度计测定间作田内外的光强。间作田光强的测点分别设在树行以东和树行以西的两个林冠下(各距树行80 cm), 另一个点设在两树行的中央。测光传感器水平置于作物冠层表面以上10 cm。对照田的测定与前者同步进行。全天从6~18时每隔2小时测定1次, 每个测点每次重复测定3次。
1.3 作物物候期、叶面积和产量测定作物出苗后选出20株进行物候观测, 每日或隔日观测1次。出苗后每15天从田间挖取作物10~20株, 洗净, 分器官烘干, 计算各时期不同器官的干重。叶面积测定是用打孔器切取叶圆片, 烘干称重, 测出比叶面积(cm2/mg), 再按叶干重计算植株的叶面积。作物成熟时按行收割, 分别统计距林木不同距离的作物产量和生物量。
1.4 光-光合速率曲线的测定和拟合在一定的物候期, 分别从田间选取生长正常的小麦、大豆和玉米植株, 在室内用FQ-WCO2分析仪, 外接056型计录仪(HITACH), 用开放气路落差法, 测定3种作物功能叶在不同光强下的光合速率。用两支500W的碘钨灯和调压变压器调节光强。所获数据用模型(1)拟合光-光合速率曲线(胡新生, 1997; Bassman, 1991)。利用模型(1)获得的大量光合速率和表观量子效率(α=C0×a), 结合实测光强、叶面积系数等参数, 计算群体光合速率(王天铎, 1987; 门司正三, 1980; 龚垒 1989)。
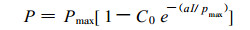
|
(1) |
P为光合速率(μmolm-2s-1); Pmax为最大光合速率(μmolm-2s-1); C0为待定参数; a为特定参数; I为光照强度(μmolm-2s-1)。
1.5 群体日累积净光合量和光能利用率的估算 1.5.1 群体各叶层表面光强的计算根据各物候期测得的间作田各点冠层表面光强、作物(或林木)的叶面积系数、群体叶幕消光系数等参数, 利用模型(2)计算出各叶层表面的光强(王天铎, 1987;门司正三, 1980; 龚垒, 1989)。
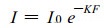
|
(2) |
I为群体某叶层下部的光强(μmolm-2s-1); I0为群体某叶层表面的光强(μmolm-2s-1); K为群体消光系数; F为群体叶面积系数。
1.5.2 群体各叶层光合速率的计算根据各叶层表面的光强, 作物的最大光合速率(Pmax)和表观量子效率(α), 利用模型(3)计算群体各叶层的光合速率(μmolm-2S-1), 将其减去叶呼吸消耗, 获得各叶层的净光合速率(4)。各叶层的净光合速率累加即为群体的净光合速率。参考有关文献(王天铎, 1987;门司正三, 1980; 龚垒, 1989; 王永锐, 1991), 玉米、小麦的消光系数(K)分别为0.50和0.60, 大豆为0.70, 741杨为0.60。
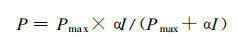
|
(3) |
P为某叶层的光合速率(μmolm-2s-1); Pmax为叶片的最大光合速率(μmolm-2s-1); J为群体某叶层表面的光强(μmolm-2s-1); α为作物或林木的表观量子效率。
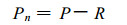
|
(4) |
Pn为叶层的净光合速率(μmolm-2s-1); R为叶层的呼吸速率(μmolm-2s-1)。
1.5.3 群体叶幕日累积光量和日累积净光合量的计算根据颜景义提出的小麦日累积光合量估算方法(颜景义, 1996), 将其进行适当改进, 提出计算群体日累积光量和日累积净光合量的模型(5);利用各时刻测得的光强和算得的群体净光合速率, 分别计算单位面积土地日累积光量和日累积净光合量。

|
(5) |
Pn (D)为日累积净光合量(m mol m-2); Pi为第i次实测叶幕净光合速率(μmol m-2s-1); Pi+1为第i+1次实测叶幕净光合速率(μmol m-2s-1); ti为第i次到第i+1次观测的时间间隔(s); n为观测次数; a为从日出到P1次测定的时间间隔(s); b为从Pn次观测到日落的时间间隔(s)。
2 结果与分析 2.1 作物的物候期和叶面积系数表 2列举了不同作物的物候期和叶面积系数(LAI)的统计结果。可以看出, 对照田作物的LAI一般都高于间作田, 其叶片衰老较慢。
|
|
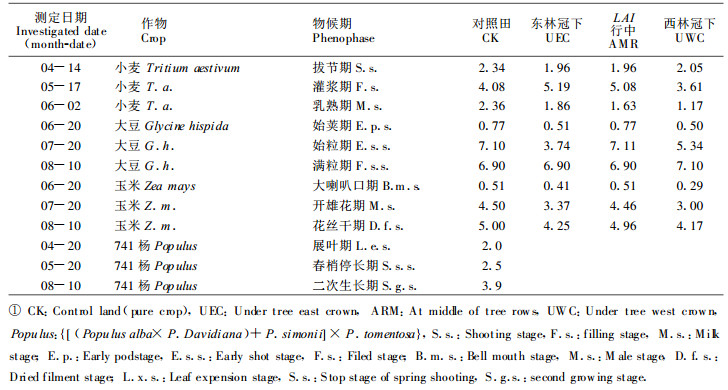
表 3列举了不同作物光-光合速率反应模型的各项参数, 其中的α=C0a为表观量子效率, LCP为光补偿点, LSP为光饱和点。从模型看出, 随生育期的进展最大净光合速率有逐渐提高的趋势。
|
|
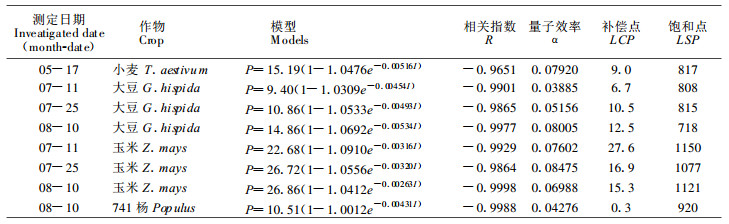
根据各物侯期间作田和对照田光强(I)日变化, 作物或林木的叶面积系数(F)、消光系数(K)、表 3提供的Pmax和表观量子效率(α), 利用模型(3)、(4)计算出群体各时刻某叶层的净光合速率(Pn)。将其相加, 获群体叶幕光合速率。3个时期的计算结果列入表 4、5。
|
|

|
|
大豆和玉米间作田因受光强度过低和呼吸消耗的影响, 叶幕净光合速率在早晚有时出现负值; 小麦受光强度较高, 没有出现这种现象。
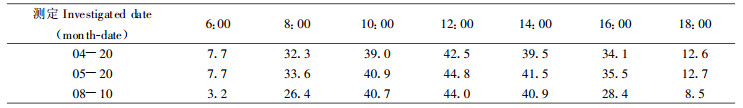
2.3.2 群体日累积光量和叶幕日累积净光合量根据1995~1996两年6个周日测定的光强日变化, 利用模型(5)计算出各测定日田间不同部位日累积光量和叶幕日累积净光含量, 结果列入表 6。日累积光量可划分为6月20日以前和以后两个阶段。可见, 在同一间作田内, 夏熟作物受林木的遮荫作用较轻; 如从4月14日到6月20日, 间作田日累积光量平均为对照田的63.95%~67.05%。秋熟作物受林木的遮荫作用较重, 如7月20日和8月10日间作田日累积光量平均为对照田的50.96%, 这是秋熟作物减产率高的重要原因。杨农间作田全年各季的遮荫程度不同, 主要是受林木叶面积生长的影响。例如, 春季741杨只有春梢生长, LAI为2.0~2.5, 夏秋季随着二次生长, LAI增加到3.9, 从而引起遮荫程度增加。
|
|
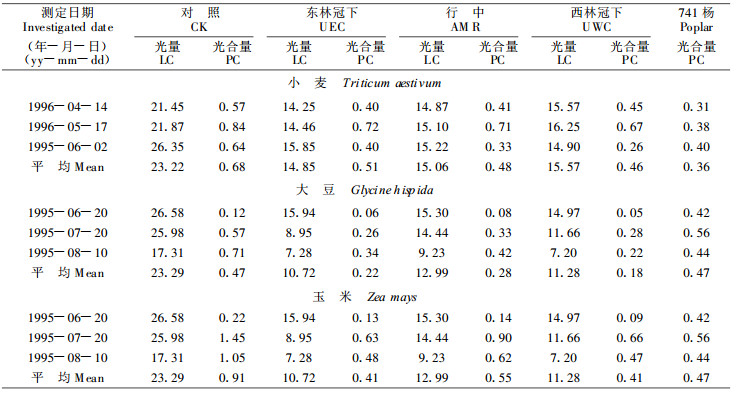
将两个林冠下的权重分别定为行中的权重定为算得间作田小麦日平均累积光量为对照的65.2%, 大豆和玉米日平均累积光量为对照的51.52%;小麦日平均累积净光合量为0.48mmolm-2, 大豆和玉米分别为0.24 mmolm-2和0.48 mmolm-2; 分别为对照田的70.59%、51.06%和52.75%。可见, 日累积光量降低一半, 日累积净光合量也约降低一半。从表 6的材料看出, 日累积光量与日累积净光合量呈简单的线性正相关。但不同作物其直线的斜率不同; 小麦、大豆和玉米的斜率分别为0.02391、0.02128和0.03973。斜率越大, 表明该种作物对光强变化越敏感。
2.3.3 群体光能利用率根据表 6提供的日平均累积光量和日平均累积净光合量, 计算各处理的光能利用率(表 7)。将间作田各部位农作物的光能利用率加权平均, 并与对照比较。可看出, 小麦的光能利用率高于对照10.89%, 间作田的大豆和玉米未能提高光能利用率。这可能是因夏作物处于更弱的光照条件下, 其光合速率降低, 但叶片的呼吸消耗速率并未降低, 导致弱光下未能提高光能利用率。
|
|

741杨的东西冠幅只占行距的一半, 故将其光能利用率折半并与间作物的光能利用率相加, 则杨-麦间作的总光能利用率为对照的135.32%, 杨-大豆和杨-玉米间作的光能利用率分别为对照的148.51%和127.62%。
农作物产量的研究表明, 小麦东、西两林冠下的平均产量为行中的86.61%;间作田加权平均产量为对照的76.09%。大豆东、西两林冠下的平均产量为行中的51.37%;间作田加权平均产量为对照的64.19%。玉米东、西两林冠下的平均产量为行中的62.37%, 间作田加权平均产量为对照的51.24%。小麦减产率最低, 玉米减产率最高。这与日累积光合量的测定结果相似。
3 结论1) 杨农间作立体种植, 冠层总叶幕系数增大; 低叶量与高叶量时期互补; 截获的光能比对照高, 从而提高了总光能利用率。2) 3种间作物的测定结果表明, 农林总光能利用率比对照提高27.56%~48.64%。某种作物在全日照下的光能利用率越低(如大豆), 间作后总光能利用率比对照提高的幅度越大。3)杨农间作当作物冠层表面的日均光强大于60%时, 间作物的光能利用率高于对照10.89%, 说明弱光下光能利率可提高。但如果光强小于50%, 则间作物的光能利用率与对照相近, 即不能提高光能利用率。这种现象可能与弱光下光合速率降低, 而呼吸消耗并未减少有关。4)杨农间作作物的日累积净光合量与日累积光量呈线性正相关, 随作物喜光程度的增加, 直线的斜率变大。
用上述方法估算的间作田与对照田农作物的日累积净光合量的比值与实测两者产量的比值基本相同说明方法的可行性。
龚垒. 1989. 树木光合作用与物质生产.树冠光合作用的数学模型(126-131). 北京: 北京科学技术出版社.
|
高学增, 高新学. 不同玉米群体CO2含量和光合作用的研究. 山东农业科学, 88(1): 19-23. |
胡新生, 等. 1997. 四个杨树无性系在不同温度和相对湿度条件下净光合速率变化的比较研究. 林业科学, 33(2): 107-116. DOI:10.3321/j.issn:1001-7488.1997.02.002 |
李明启. 1980. 关于植物光能利用效率与作物产量问题.光合作用研究进展,. 北京:科学出版社: 171-178. |
刘建栋, 傅抱璞. 1996. 光胁地条件下油菜产量及其生产潜力的研究. 中国农业气象, 17(2): 22-25. |
孟平, 宋兆民, 张劲松. 1997. 农林复合系统光能利用率的研究. 林业科学, 33(专刊1): 14-19. |
门司正三左伯敏郎.植物群体中光因素及其对植物生产的作用.光合作用与作物生产译丛(2), 北京: 农业出版社, 1980: 1~ 24
|
裴保华, 蒋湘宁, 郑均宝, 等. 1990. I -69杨光能利用的研究. 林业科学研究, 3(3): 201-206. |
王天铎. 1987. 光合作用生理生态若干问题.植物生理学专题讲座———纪念罗宗洛教授. 北京: 科学出版社, 97-104.
|
王永锐. 1991. 作物高产生理. 科学技术出版社: 33-48. |
颜景义, 郑有飞, 韩素芹. 1996. 小麦日累积光合量估算方法探讨. 中国农业气象, 17(5): 23-27. |
杨敏生, 林青, 邢秀岩. 1991. 741杨光合性能的研究. 河北林学院学报, 6(3): 161-165. |
张祝平, 丁明懋. 1996. 鼎湖山亚热带季风常绿阔叶林的生物量和光能利用效率. 生态学报, 16(5): 525-534. |
Johon H.Bassman, Johon C.Zw ier. 1991. Gas exchange characteristic of Populus trichocarpa, Populus deltoides and Populus trichocarpa ×Populus deltoides clone. Tree Physiology, 8: 145-159. DOI:10.1093/treephys/8.2.145 |