 2000, Vol. 36
2000, Vol. 36文章信息
- 洪伟, 吴承桢, 林成来, 赖建明.
- Hong Wei, Wu Chengzhen, Lin Chenglai, Lai Jianming.
- 福建龙栖山森林群落林窗边缘效应研究
- GAP EDGE EFFECT OF THE FOREST COMMUNITIES IN LONGXI MOUNTAIN, FUJIAN PROVINCE
- 林业科学, 2000, 36(2): 33-38.
- Scientia Silvae Sinicae, 2000, 36(2): 33-38.
-
文章历史
- 收稿日期:1998-10-13
-
作者相关文章


森林林窗(Forest gap)是由某一林层树木死亡等原因造成林地上形成不连续的林中隙地, 它是森林更新和生长的潜在空间。该空间特殊部位即林窗与森林的结合部位构成林窗边缘, 由于环境梯度变化和各植物物种在发生、竞争等过程上差异, 出现明显边缘效应。森林边缘效应对森林物种多样性的维持和更新过程将产生重大的影响, 实际上成熟的森林生态系统就是无数林窗较为固定动态平衡的反映, 因此该领域的研究不但具有生态学意义, 而且对森林的动态管理也具有现实意义。
70年代以来, 林窗及边缘效应研究成为森林动态研究的重点(王伯荪, 1986; 牛文元, 1990; 马世骏, 1990; 奚为民, 1993; Brokaw, 1985; Hybbel, et al.1986; Thomass, 1985; 奚为民, 1992)。对于边缘效应, 国内王伯荪(1986)研究了森林群落边缘效应; 牛文元(1990)提出了生态环境脆弱度指标及脆弱带的函数表达; 马世骏(1990)提出了边缘效应的概念及其研究的生态学意义; 奚为民(1993)研究了四川缙云山森林群落林窗边缘效应。本文对福建龙栖山国家级自然保护区森林群落林窗测量其边缘效应强度,探讨林窗边缘的特征和性质, 为森林的营造和优化管理提供科学依据。
1 研究地自然概况龙栖山自然保护区地处福建省西北部, 东经117°11′~117°21′、北纬26°23′~26°43′, 南北长6.5km, 东西宽7 km, 总面积126 km2。主峰海拔高1620.4 m, 海拔1000 m以上的山峰有14座, 土壤母质主要由花岗岩、变质岩、砂砾岩、云母片岩等组成; 一般在海拔800 m以下主要为红壤与黄壤, 800 m以上为黄壤或粗骨性黄壤, 而山顶主要为山地草甸土。该区山体崎岖, 溪流比较大, 流速较快, 水质尚好, 水热资源丰富, 属亚热带季风气候, 多东北风, 其次为西南风。根据保护区山下的将乐县气象站的资料表明:该地区年均气温14~19℃, 1月最冷平均气温8.0℃, 7月最热平均气温23℃; 年平均降雨量为1600~1800 mm; 年均相对湿度82.0%, 年日照时数1736 h, 无霜期300~339 d, 每年降雨约5~6次。
龙栖山保护区山下人口密集, 林木破坏比较严重, 但山里居住人口较少, 植被保存尚好, 山地有成片常绿阔叶林是其主要植被, 一般生长在1300 m以下常有栎属(Quercus)、栲属(Castanopsis)、樟属(Cinnamomum)等乔木为建群树种。落叶阔叶林中乔木一般有长柄水青冈(Fagus longipetiolata)、缺萼枫香(Liquidambar acalycina)、桦属(Betula)、椴属(Tilia)等组成。
2 研究方法 2.1 取样数量以对森林群落全面踏查为基础, 对针阔混交林、常绿阔叶林、常绿落叶阔叶林群落各选择9个林窗样地, 其中针阔混交林发育前、中、后期各3个, 共调查林窗样地27块, 取样总面积1396 m2。
2.2 发育期的成分林窗发育期的划分主要依据林窗形成时间, 并参考林窗内植被高度和物种结构确定。前期林窗形成时间短, 林窗内植被以少数几种喜阳草本占优势, 如里白(Diplopterygium glaucum)、芒萁(Dicranopteris pedata)等, 且盖度很大, 一般高度低于2 m。中期林窗边缘的喜阳树种的树冠有明显的向林窗内倾趋势, 林窗内植被以木本为主, 里白大量死亡, 林窗内小乔木一般高度低于8 m。后期林窗已进入发育晚期, 林窗中心的植物种与其边缘区基本一致, 林窗内植被高度已接近原群落树冠高度, 而林下则以苗木及耐荫草本为主。
2.3 取样方法采用样方法取样, 对所确定的每个林窗记载其环境特征和周围群落性质, 对构成林窗边界的各大乔木进行每木调查; 以林窗边界乔木树干之间的距离为准, 采用矩形法确定林窗大小。在每个林窗中心区、边缘区以及非林窗地段设置3个4 m×4 m的样方, 分别记载其中的小乔木和灌木的种类、个体数、高度、冠幅等值; 对样方中的草本植物及林灌幼苗记录其种类、个体数和盖度等内容。
2.4 测定方法采用Shannon-Wiener指数测定林窗不同区域的物种多样性(D)
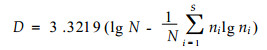
|
(1) |
式中, N为所有种的个体数; ni为第i种的个体数; S为物种数。
以Simpson指数测定林窗各区域的生态优势度(C)
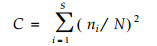
|
(2) |
式中, N、S和ni的含义与(1)式相同。在此基础上, 以王伯荪等确定的边缘效应指数测定林窗的边缘效应强度(E) (王伯荪, 1986) :
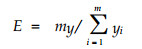
|
(3) |
式中, y为m个群落形成的交错区里度量群落内种群数量和结构的定量指标。本文选用ShannonWiener指数和Simpson指数分别计算(3)式, 得出分别由D值和C值计算出的边缘效应值ED和EC:
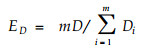
|
(4) |
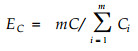
|
(5) |
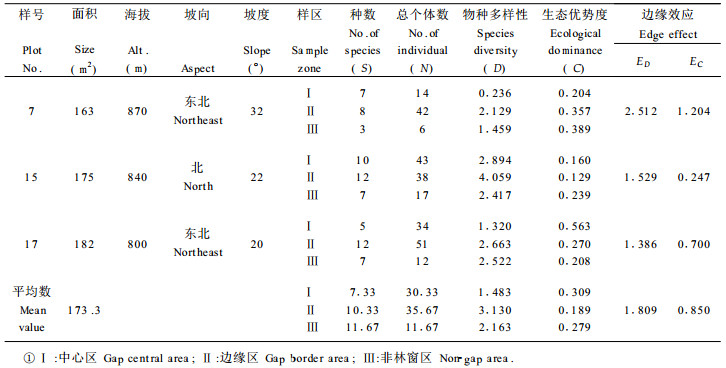
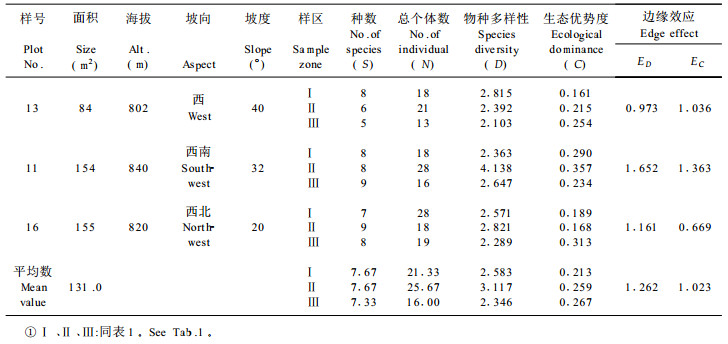
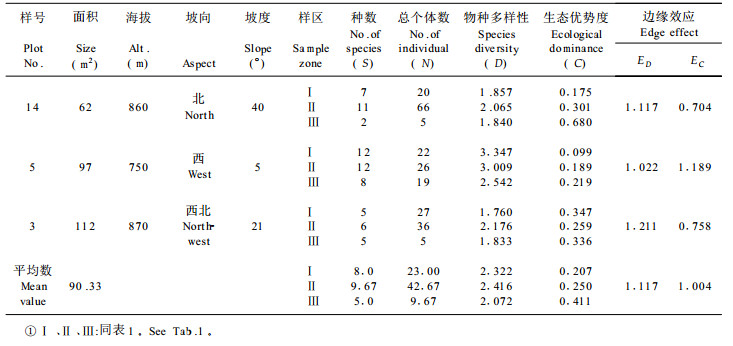
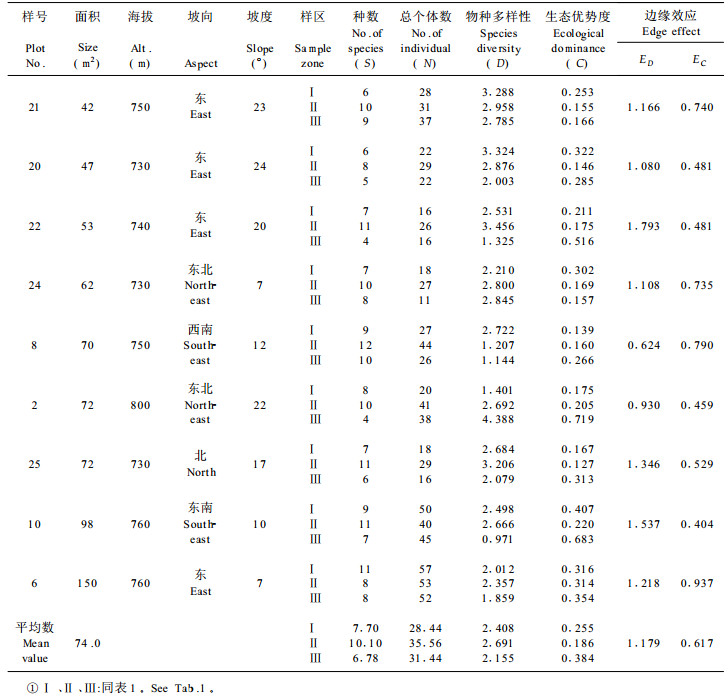
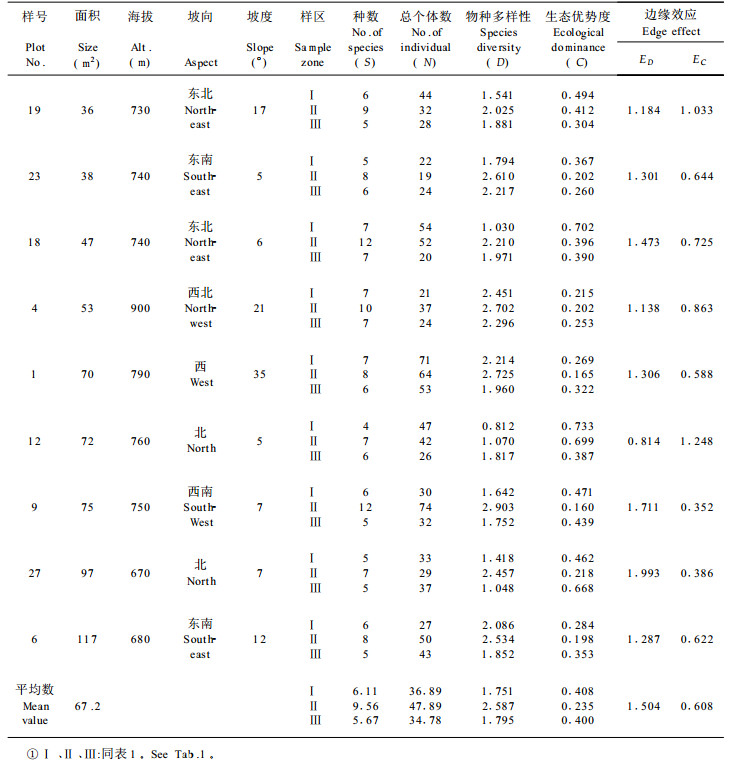
根据27个林窗样地的取样数据, 由(1)、(2)式计算出各林窗中心区、边缘区和非林窗地段的物种多样性指数和生态优势度指标, 再由(4)、(5)两式分别求出各林窗相应的边缘强度值, 其计算结果按各林窗所处的不同森林类型、发育阶段和林窗面积的大小分别列于表 1至表 5。
|
|
|
|
|
|
|
|
|
|
从物种构成上看, 龙栖山自然保护区针阔混交林中林窗中心区常见的植物多为喜阳植物, 如里白(Diplopterygium glaucum)、芒萁(Dicranopteris pedata)、箬竹(Indocalamus nakai)等, 它们构成了森林群落更新的先锋植物, 而在林窗边缘区则为多种植物生态型物种的汇合, 如罗浮栲(Castanopsis fabri)、鹿角杜鹃(Rhododendron rarrerae)、绒毛润楠(Machilus velutina)等常见植物。林窗边缘区的植物种数和总体数均大于林窗中心区, 且结构也较复杂(表 1~3)。常绿阔叶林、常绿落叶混交林均存在与针阔混交林中林窗相类似的特征, 只是物种有所不同而已, 如常绿阔叶林林窗常见鳞果椴(Tilia endochrysea)、乌药(Linderra aggregata)、梅叶冬青(Ilex asprella)等物种, 而常绿落叶混交林林窗中常见物种有山姜(Alpinia japonica)、
同一林窗样地不同区域物种多样性测定结果, 都充分显示林窗边缘区由于边缘效应的作用有增大物种多样性的趋势(表 1~5), 其中以针阔混交林中前期林窗的这种趋势最为明显(表 1), 其次为常绿落叶混交林的林窗(表 5)。生态优势度的变化趋势表现了与物种多样性呈负相关的规律, 即所有林窗样地的中心区都表现为较小的物种多样性值和较大的生态优势度值, 而在林窗边缘区上述两种数量指标则相反。例如, 第17号林窗样地中心区的样方植物种数为5, 总个体数为34, 物种多样性指数为1.320, 生态优势度为0.563;而该林窗边缘样方植物种数为12, 总个体数为51, 物种多样性系数为2.663, 生态优势度为0.270;两者物种多样性指数之差为1.343, 生态优势度之差为0.293 (表 1)。
林窗所有特征中最重要的属性是动态特征。不同发育阶段的林窗的物种多样性、生态优势度和边缘效应强度的测定揭示了随时间的推移林窗组成结构的动态趋势。计算结果充分说明林窗从前期到发育的中、后期,林窗中心的物种多样性指数有增大趋势,生态优势度的变化则与之相反。这种变化形成了相应的林窗边缘效应强度动态, 即随着林窗由前期向中、后期发育, 以物种多样性为基础得出的林窗边缘效应强度值ED逐渐减小, 而以生态优势度求算出的林窗边缘并行应强度EC却有增大的趋势, 这充分反映了随着林窗的不断发育, 林窗趋于向原森林群落恢复, 边缘效应逐渐消失的动态规律。
常绿阔叶林、常绿落叶混交林的林窗组成结构数量特征及其边缘效应变化规律与针阔混交林基本一致(表 1~5), 但由于森林群落的不同, 导致了它们之间存在着一定的差异。从分析对比可知, 针阔混交林前期的边缘效应强度均值ED (1.809)大于常绿落叶混交林的林窗边缘效应均值ED (1.504);而常绿落叶阔叶林之边缘效应强度均值又大于常绿阔叶林的均值ED (1.345), 这说明针阔混交林的林窗边缘效应强度在一定意义上大于阔叶林林窗之边缘效应。
从这次调查结果来看, 林窗的面积变化幅度不是很大, 27个林窗的面积基本上在40~190 m2之间, 其中针阔混交林前期林窗平均面积为173.3 m2, 中期林窗的平均面积为131.0 m2, 后期林窗平均大小为90.3 m2, 这说明随着林窗的发育, 处于同一森林类型的林窗面积逐渐减小。林窗的大小影响着林木的更新,而更新又影响冠层的树种组成,因此林冠的树种组成与林窗的大小之间存在某种相关关系。林窗的大小不仅在很大程度上决定了林窗的环境特征, 如光照、温度和湿度等的波动变化, 而且影响着林窗植被的物种多样性和边缘效应的强度, 从而构成影响林窗植被更新的重要方面。
4 讨论森林群落林窗的中心到森林群落内部光、热、水等环境因素存在显著梯度变化, 这使得森林内部构成一定的生态分异。森林植物生态学性质的差异导致林窗内部, 特别是在林窗边缘区发生较为频繁的种间和种内竞争过程。林窗的不断形成、边缘效应机制的制约, 使得林窗成为保护和恢复森林群落物种多样性的重要场所。龙栖山森林群落林窗边缘效应研究表明, 林窗边缘区具有增大物种多样性的趋势, 因此, 可以认为林窗边缘效应属于具有动态特点的正效应, 从林窗的形成、发育到消失, 林窗边缘效应的强度随着森林的更新发生相应变化, 它反映和指示了森林的更新性质。
龙栖山森林群落林窗边缘效应的研究, 可以为混交林的种间结构重建以及选择最佳林木择伐面积和强度, 以利于森林物种多样性的保持和林木更新提供科学依据。林窗边缘效应与林窗面积、坡度、坡向等环境因子之间的关系还有待于深入研究,以不断丰富和发展林窗理论。
马世骏.边缘效应与边际生态学.见:现代生态学透视.北京:科学出版社, 1990, , 43~45
|
牛文元.生态环境脆弱带的基础判定.见:现代生态学透视.北京:科学出版社, 1990, 46~53
|
王伯荪. 1986. 鼎湖山森林群落分析X.边缘效应. 中山大学学报(自然科学版), 25(4): 52-56. |
奚为民, 钟章成, 毕润成. 1993. 四川缙云山森林群落林窗边缘效应的研究. 植物生态学与地植物学学报, 17(3): 232-242. DOI:10.3321/j.issn:1005-264X.1993.03.005 |
奚为民. 1992. 林窗植被研究进展. 西南师范大学学报(自然科学版), 17(2): 268-274. |
Brokaw N V L. 1985. Gaps phase regeneration in tropical forest. Ecol, (66): 682-687. |
Hybbel S P, Foster R B. Canopy gaps and the dynamics of a netropical forest.From " Plant Ecology", Chapter 3(Ed. by Craw ley, M. J.), Blackw ell scien tific publication, 1986, 77~96
|
Thomass V T. 1985. Forest development in tree-fall gaps in the tem perate rain forests of Chile. Natl. Geogr. Res., 1(2): 162-183. |