 2000, Vol. 36
2000, Vol. 36文章信息
- 郑均宝, 梁海永, 田颖川, 高宝嘉, 王永芳.
- Zheng Junbao, Liang Haiyong, Tian Yingchuan, Gao Baojia, Wang Yongfang.
- 转双抗虫基因741毛白杨的选择及抗虫性
- SELECTION AND INSECT RESISTANCE OF TRANSGENIC HYBRID POPLAR 741 CARRYING TWO INSECT-RESISTANT GENES
- 林业科学, 2000, 36(2): 13-19.
- Scientia Silvae Sinicae, 2000, 36(2): 13-19.
-
文章历史
- 收稿日期:1999-11-10
-
作者相关文章


2. 中国科学院微生物研究所 北京 100080
2. Institute of Microbiology, Academia sinica Beijing 100080
虫害是造成农、林业减产的重要原因之一。半个世纪以来, 人们使用化学农药、微生物杀虫剂等方法防治害虫, 曾起过很大的作用, 但也出现了不少意想不到的问题。现在分子生物学, 特别是80年代成熟起来的植物基因工程, 为防治害虫提供了一条新的途径(朱新生, 1997)。Mc Comn(1991)首次报道了获得一株有杀虫效果的杨树转Bt基因植株。我国也在1993年报道了获得一批转Bt CrylAc基因的欧洲黑杨, 对舞毒蛾和杨尺蠖毒杀死亡率可达到80 %~ 90 %(田颖川, 1993)。Confalonieri M.等(1998), 报道了用大豆蛋白酶抑制剂基因转化了欧洲黑杨, 但对试虫的毒杀效果并不好。近年的研究指出, 昆虫易对B-to xin产生耐受性, 并认为防止害虫耐受性发生最有效的方法是联合使用两种或两种以上不同杀虫机制的抗虫基因, 使转基因植物表达数个具有不同毒性机制的毒素(朱新生, 1997)。田颖川等对Bt CrylAc基因部分片断的核苷酸序列进行了改造与慈菇蛋白酶抑制剂(API)基因构建了具有不同杀虫机制的双抗虫基因, 并成功的转化了741毛白杨[Populus alba L.×(P.davidiana Dode + P.simonii Carr.)×P.tomentosa Carr.], 经过抗虫试验及分子生物学检测在国内、外首次报道选到了对杨扇舟蛾和舞毒蛾有高抗虫性的转双抗虫基因杨树植株。本文重点报道对转双抗虫基因741毛白杨的选择条件, 选出一批高抗虫性和中等抗虫性植株(无性系), 其方法也可供其它类似研究的参考。
1 材料和方法 1.1 材料 1.1.1 菌种和质粒含双抗虫基因的表达载体pBtiA, 及土壤农杆菌LBA4404(田颖川, 2000)。
1.1.2 植物材料和测试昆虫741毛白杨取自河北农业大学林业生物技术研究室无菌试管植株。测试昆虫为人工饲养的杨扇舟蛾(Clostera anachoreta Fabricius)和舞毒蛾(Lymantria dispar Linnacus)。
1.2 方法 1.2.1 双抗虫基因的构建含部分改造的Bt CrylAc基因(Bt基因)及慈菇蛋白酶抑制剂基因A (API—A)的植物表达载体的构建(田颖川, 2000)。在转化植物之前将含有该载体的土壤农杆菌LBA4404, 在28 ℃摇床培养过夜后即可用于转化。
1.2.2 双抗虫基因对杨树的转化及外植体诱导器官分化抗卡那霉素(Km')临界浓度试验以试管内杨叶片为外植体, 按附后文献(田颖川, 1993)的方法进行转化。诱导不定芽分化的培养基为MS +6 - BA 1.0 mg/L +NAA 0.1 mg/L; 诱导芽增殖培养基为MS +6 -BA 0.3 mg/L +IBA 0.1 mg/L; 诱导嫩茎生根的培养基为1/2MS +IB A0.3 mg/L(或NAA 0.1 mg/L)。抗Km'临界浓度试验是在上述相应培养基中附加Km'浓度分别为0、5、10、20、30、40、50、100 mg/L等。诱导嫩茎生根抗Km'临界浓度试验, Km'的浓度分别为0、10、20、30、40、50 mg/L。
1.2.3 转基因植株的饲虫试验饲虫材料为苗圃试验地上的转基因和未转基因(CK)的741毛白杨各系号苗木的叶片, 进行室内饲养。饲养瓶为高6 cm直径12 cm的罐头瓶, 用塑料薄膜扎紧瓶口, 细针扎孔通气。选取孵化1~2 d的一龄杨扇舟蛾和舞毒蛾开始饲养, 每天更换新鲜叶片, 直到结茧为止。隔日观察一次, 记载昆虫死亡数, 死、活虫发育的龄级, 定期称取虫重等。在1998年和1999年进行了两年饲虫试验, 参试植株21个系号。由于CK植株昆虫死亡率都在5 %以下, 因此只计算昆虫死亡率, 免去校正死亡率的计算。计算饲虫过程中蜕皮指数和毒力(毒效)的公式如下:
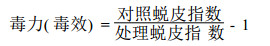
|
数值愈高说明毒力愈大。
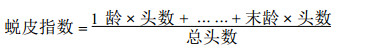
|
选择抗虫性强、中等、无显著抗虫性的转基因系号, 以及未转基因(CK)等的嫩枝扦插小苗高20 cm左右, 按随机区组设计, 栽植在苗圃试验地上, 共设4个小区, 每个系号共栽40株。苗木生长了120天, 高生长停止后, 逐株测定苗高。各系号选出最大值的5株, 求其平均值(X)和标准差(S), 进行t检验。同时观察生长正常的一年生和两年生苗木的干形、分枝角、叶片形态等。
1.2.5 转化再生植株的分子生物检测杨树DNA的提取 PCR检测及DNA Southern杂交分析参照附后参考文献(Ausubel, 1995)等进行。将20μ g杨树叶总DNA, 用Hind Ⅲ酶切过夜后用于Sothernblot杂交分析, 探针分别为32P标记的Bt基因(1.8Kb)和AHPI基因(0.6Kb)片段。
杨树叶蛋白的提取及免疫测定 蛋白质的提取按照附后参考文献中Perlak FJ等(1991)所述的方法进行。在ELISA分析中, 首先用1: 2500稀释的Bt蛋白鸡抗4 ℃包被过夜, 然后加已知浓度的标准抗原或植物蛋白溶液37 ℃反应3 h, 再加入1: 1000稀释的Bt蛋白鼠抗37 ℃反应2 h, 最后加入羊抗鼠酶标二抗37 ℃反应2 h, 每步均用PBST洗板3次。最后加入底物避光反应半小时, 用BioRad 550型酶标仪测定结果。
2 结果与分析 2.1 不同器官抗Km'临界浓度的确定及转化基因植株的初步选择 2.1.1 不同器官抗Km'临界浓度的确定由于所用菌种带有抗Km'基因(NPTII, 新霉素磷酸转移酶标志基因), 如基因已转入杨树并得到表达, 该植株在含有一定浓度Km'培养基上, 能正常生长。根据试验, 外植体的不同器官对Km'的耐受能力不同, 为此必须在试管内进行叶片诱导不定芽发生、嫩茎的增殖、嫩茎的生根等抗Km'临界浓度试验。
试验结果说明:叶片培养了42 d, 嫩茎培养了46 d, 培养基Km'从0到50 mg/L呈梯度增加, 引起的变化为叶片变黄进一步坏死, 以及不定芽发生或芽的增殖由多到少, 到完全不发生等; 嫩茎在含有浓度为20 mg/L Km'培养基中, 上部叶绿, 下部叶黄, 直到整株变黄, 稍有绿色, 芽的增殖也是由多到少。确定诱导叶片不定芽发生的抗Km'临界浓度为30 mg/L, 在此培养基中培养, 叶片变黄, 但尚未坏死, 没有不定芽发生。而诱导嫩茎芽增殖的临界浓度为50 m g/L, 在这种培养基中, 嫩茎整株变黄, 稍有绿色。试验结果还说明Km'对诱导嫩茎生根具有更大的抑制作用, 浓度为10 m g/L即可达到完全抑制生根的效果。而转基因植株即使Km'浓度达到50 mg/L, 仍能正常生根。
2.1.2 转化基因植株的初步选择以试管内无菌叶片为外植体, 按照田颖川等(1993)的方法进行基因转化。叶片经过浸菌和共培养后, 放置于附加30 mg/L Km'和200 mg/L的羧苄青霉素的诱导不定芽分化的MS培养基中(见1.2.2), 大约培养20~30 d, 叶片开始大部分坏死, 呈水渍状但在切口部位及叶柄剪口处出现米粒状愈伤组织, 经进一步培养, 诱导出不定芽, 约40~50 d形成丛状芽丛。培养过程中并不是所有叶片均能产生不定芽, 有些虽形成了绿色愈伤组织, 但未能继续分化形成不定芽。能够诱导产生不定芽的叶片仅占培养叶片总数的2.5 %左右。
抗Km'临界浓度的试验说明, 嫩茎不定根的分化对Km'更加敏感, 因此对培养的抗Km'不定芽先不进行增殖培养, 而是在生根培养基中选择抗Km'的生根植株。将抗Km'的不定芽培养为1.5~2.0 m高的嫩茎, 转到含30 mg/L Km'和100 mg/L的羧苄青霉素的生根培养基中, 约有50 %左右的植株能生根, 其余不能生根的植株认为是没有转化基因的。在试管中进一步繁殖这些初步选择的转基因植株, 形成无性系, 加以编号, 并栽植在苗圃中的试验地上。对初步选择的转基因植株, 随机抽取10个系号进行PCR检测, 80 %呈现阳性反应。
2.2 具抗虫性转基因植株的饲虫试验选择由于对舞毒蛾已有报道(田颖川, 1993), 而且所得抗虫杨树对杨扇舟蛾的抗性较低, 所以本文重点介绍杨扇舟蛾的饲虫试验。
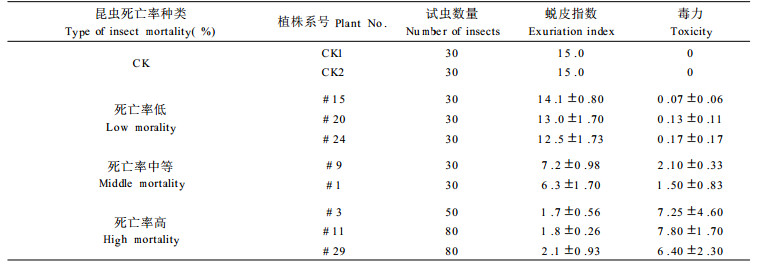
2.2.1 杨扇舟蛾的死亡率和定期死亡率饲虫试验从孵化1~2 d的幼虫开始, 昆虫经过5个龄级的发育到作茧结束, 历时23~25 d。统计各系号植株的杨扇舟蛾死亡率, 即试验全过程死亡的虫数占饲虫总数的百分率, 绘入图 1中。图 1表明: (1)未转基因的对照昆虫死亡率极低, 两次均为3.4 %。(2)昆虫死亡率低, 为20 %以下, 占参试系号的61.9 %。如图 1中的#24、#20和#15。(3)昆虫死亡率中等, 为40 %~ 70 %, 如图中的#1为65 %±7.6 %, #9为53.3 %±5.8 %, #10为46.4 %±11.5 %, #12和#17为60.0 %±10.0 %, 占参试植株的23.8 %。饲养舞毒蛾, #1植株昆虫死亡率为66.3 %± 0. 4 %。(4)昆虫死亡率高, 达85 %以上, 如图中的#3为89.6 %±5.2 %, #11为88.4 %±5.9 %, #29为84.6 %±6.0 %, 占供试植株的14.3 %。饲养舞毒蛾, #3植株死亡率可达91.8 %±11.7 %, #11为90.5 %±0.7 %。昆虫定期死亡率, 指在昆虫饲养的过程中, 一定时间内(每2 d内), 死亡昆虫数占死亡昆虫和活虫之和的百分率。计算各系号植株的昆虫定期死亡率, 作图 2。图 2说明: (1) CK的昆虫死亡率低, 其定期死亡率也低, 昆虫死亡发生在饲养初期。(2)昆虫死亡率低的植株, 如#20, 第3 d和第5 d定期死亡率分别为6.7 %和7.1 %, 与CK相似。或出现昆虫偶尔死亡情况, 如植株#15, 在第9 d定期死亡率为5 %。(3)昆虫死亡率中等的植株, 定期死亡率较低, 其延续时间长。(4)昆虫死亡率高的植株, 定期昆虫死亡率较高, 而且主要是集中在饲养初期。

|
图 1 昆虫死亡率 Fig. 1 Mortality percentage of larvae |

|
图 2 饲虫过程中昆虫定期死亡率 Fig. 2 Mortality percentage of larvae in fixed peried during feeding (1)对照Control, (2)死亡率低Low mortality, (3)中等死亡率Middle mortality, (4)高死亡率High mortality. |
饲养过程中, 除死亡的昆虫外, 还存活一定数量的昆虫。转基因植株体内所含有的毒蛋白或蛋白酶抑制物质对它们的发育有什么影响?为此研究了对这些活虫的发育龄期、化蛹期长短以及虫体重量的影响, 其结果列入表 3和表 4中。表 3表明, 转基因植株与对照相比, 死亡率中等和死亡率高的植株, 其叶片饲虫的结果, 会推迟存活昆虫的发育龄期, 从Ⅰ龄进入Ⅱ龄期, 昆虫中等死亡率推迟2~6 d, 高死亡率推迟3~8 d; Ⅱ龄进入Ⅲ龄为2~4 d和2~6 d; Ⅲ龄进入Ⅳ龄均为2~6 d; Ⅳ龄进入Ⅴ龄为2~8 d和4~6 d, #11植株甚至使存活昆虫不能达到Ⅴ龄、不生长、不蜕皮, 延续19日死亡。见表 1。
|
|
|
|

|
|

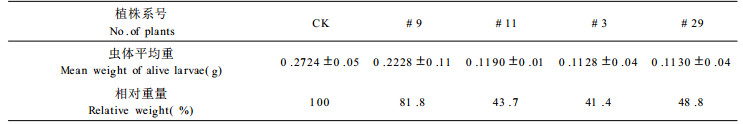
表 2表明, 昆虫死亡率高的植株, 喂养12 d, 每只虫体平均重只有对照的41 %~ 49 %, 昆虫死亡率中等的, 如#9植株为81.8 %。
|
|
根据杨扇舟蛾蜕皮指数计算毒力(毒效), 也能反映用转基因杨树叶片饲养后, 对尚存活昆虫生长发育的延迟的后效应, 毒力数愈高说明毒力愈大(田颖川, 1993)。测定了4类昆虫死亡率植株的毒力, 列入表 3中。
表 3表明昆虫死亡率中等的植株, 其毒力在1.5~2.5之间, 昆虫死亡率高者, 毒力达6~8之间。昆虫死亡率低者, 其毒力很低, 在0.05~0.20之间。
从以上饲虫试验结果, 根据昆虫死亡率和昆虫定期死亡率及致使杨扇舟蛾的发育龄期延迟以及昆虫体重降低, 毒力数字大等指标, 可选择昆虫死亡率高及昆虫死亡率中等的两类植株, 它们是具有显著抗虫性转基因的741毛白杨。
2.2.4 转基因植株苗期高生长与形态观察将苗木高生长测定和数据的数理统计处理结果列入表 4中。表 4表明, 对照生长好的植株平均高生长为180.0 cm。昆虫死亡率高和中等的两类, #3稍低于CK, #1稍同于对照。从1和2年生苗木的叶形、干形、分枝角等观察与CK相比没有显著差异。
2.3 转基因抗虫杨树植株的分子生物学检测选择昆虫死亡率高和中等的3个系号植株叶片, 即#11、#29、和#1进行DNA的Southern blot分析。将杨树DNA用Hind Ⅲ完全酶解后, 每一样品分两部分按1.2.7中所述进行电泳分离和转膜然后分别与Bt基因及AHPI基因探针进行杂交。杂交膜的放射自显影结果分别如图 3所示, 从图 2结果显示3个抗虫植株#1、#11、#29都可与Bt基因探针杂交形成大于14Kb(图 3—A泳道P)单一杂交带, 而非转基因对照则无杂交带出现。由于Hind Ⅲ只在Bt基因的上游有识别位点所以杂交带的数目可以反映该基因在杨树基因组中插入的拷贝数。因此可以认为在这3株植株中各有一个拷贝的插入。在用AHPI—A基因为探针的杂交中(见图 3—B)同样非转基因对照无杂交带, 而3个抗虫植株的DNA在1.6Kb左右有明显的杂交带出现, 证明AHPI—A基因在这3株植物的基因组中都有整合, 由于在AHPI基因的上下游都有Hind Ⅲ位点的存在所以杂交带的大小都是一致的, 这也说明该基因在这3个转基因植物中是完整的, 没有出现缺失和重组现象。以上结果表明这3个抗虫植株是整合有双抗虫基因的转基因植株。

|
图 3 再生杨树植株DNA的Southern印渍分析 Fig. 3 Southern blot analysis of genomic DNAs from insect-resistant poplar plants |
抗虫蛋白在转基因植株中是否有合成应是说明这些植株抗虫原因的直接证据。为此用抗Bt杀虫蛋白N端68KD的抗血清对转基因杨树叶片蛋白提取物进行了免疫测定, 测定结果初步证明在这3株抗虫杨树中有Bt杀虫蛋白的合成。用Bt抗血清进行的ELISA检测结果, 这3株转基因植株#1、#11、#29中Bt蛋白的表达量分别为0.0115 %, 0.0160 %和0.0125 %。
3 结论与讨论分别对741毛白杨无菌组培植株诱导器官再生的叶片、嫩茎等进行抗Km'临界浓度试验。以嫩茎诱导不定根发生对Km'最为敏感, 临界浓度在10 mg/L以下。在含Km'生根培养基中初步选择转化了的植株, 即生根的植株。PCR检测结果其中80 %以上的植株呈现阳性反应, 进行饲虫试验有38.1 %表现了明显的抗虫性。用农杆菌介导法转化基因时, 如果其标志基因为抗Km'基因, 对不同树种、不同器官以致继代多次逐渐幼化了的组培材料等均要再次进行抗Km'临界浓度试验, 这是基因转化能否成功的关键因素之一。
饲虫试验的结果, 根据昆虫死亡率, 可将参试植株分为4类: (1)未转基因植株(CK), 昆虫死亡率5 %以下; (2)昆虫死亡率低, 20 %以下; (3)昆虫死亡率中等, 30 %~ 70 %; (4)昆虫死亡率高, 71 %以上。比较了上述4类植株叶片毒死昆虫的昆虫定期死亡率。(1)和(2)两类, 昆虫定期死亡率低且发生在饲虫初期, 其特点两类近似, (2)类可判断为基因未转入, 或转入后基因未表达。第(3)类, 昆虫定期死亡率较低, 但延续时间长, 是否可解释为基因产物表达水平低, 在昆虫体内抗虫蛋白积累到一定水平才能使昆虫致死。可见第(3)类也是被选择的植株。而第(4)类, 定期昆虫死亡率较高, 而且集中在饲养初期, 说明基因表达产物水平高, 因而较迅速地毒死了昆虫。总之, 要选择第(4)类和第(3)类植株。
根据试虫蜕皮指数计算毒力, 说明叶片毒性对尚存活昆虫生长发育的延迟性后效应, 数值愈高说明毒力愈大。本研究除根据蜕皮指数计算了毒力外, 还研究了用上述4类植株叶片饲养过的尚存活昆虫各发育龄期推迟的程度。未转基因的对照和昆虫死亡率低的两类尚存活昆虫进入各发育龄期所需的时间相近; 而昆虫死亡率高和中等的两类尚存活昆虫进入各发育龄期均推迟, 有的昆虫甚至不能达到Ⅴ龄期就死亡了。可见这些尚存活的昆虫也受到毒害, 影响了正常的生长发育。但是它们是否能正常地结茧、羽化、产卵、孵化幼虫以致下一代对转基因杨树的反应等, 均有待进一步研究。
此外, 第(4)类和第(3)类喂养12 d后, 尚存活昆虫平均虫体重的显著减轻, 也说明了对昆虫生长发育的延迟性后效应。

|
图版Ⅰ Plate Ⅰ 1 叶片产生不定芽; 2 抗Km'临界浓度试验(从左到右Km'浓度0, 5, 50 mg/L); 3 在含有Km'的培养基中转基因植株生了根(右), 未转基因的未生根(左); 4 在苗圃试验地上的转基因植株; 5 在瓶内饲养1d的杨扇舟蛾的叶片; 6 在瓶内饲养42 d的存活下来的舞毒蛾幼虫(从左到右为#3, #11, #29, #15以及对照) 1 Clusters of adventitious shoots. 2 Test on the critical concentration of Km'. (from left to right 0, 5, and 50 mg/L). 3 Rooting of the transgenitic shoots cultured on the medium containing Km' (right), untransgenitic shoots without rooting(left). 4 Transgenitic plants grown in nursery. 5. The leave of feeding clostera anachoreta larvae after 1 day. 6 The alive larvae of Lymantria despair feeding 42 days(from left to right, #3, #11, #29, #15 and CK). |
田颖川, 韩一凡, 等. 1993. 抗虫转基因欧洲黑杨的培育. 生物工程学报, 9(4): 291-297. DOI:10.3321/j.issn:1000-3061.1993.04.017 |
田颖川, 虞红梅, 郑均宝, 等. 2000. Studies of transgenic Hybrid poplar 741 transformed with tw o insects resistantgenes. 植物学报, 42(3): 263-268. |
朱新生, 朱玉贤. 1997. 抗虫植物基因工程研究进展. 植物学报, 39(3): 282-288. |
Ausubel FM, Brent R, Kingston RE et al.Short protocols in Molecular biology.Third edition, USA: John Wiley & Sons, Inc.1995.2.1~2.12; 10.1~11.19
|
Confalonieri M, Allegro G, Balestrazzi A, et al. 1998. Regeneration of Populus nigra transgenic plants expressing a Kunitz proteinase inhibitor (Kti3) gene. Mol Breeding, 4: 137-145. DOI:10.1023/A:1009640204314 |
Mc Cown B H, Mc Caba DE, Russd DR, et al. 1991. Stable Transformation of Populus and incorporation of pest resistance by electric discharge particle acceleration. Plant cell Rep, 9: 590-595. |
Perlak FJ, Deaton RW, Armstrong TA, et al. 1990. Insect resistant cotton plants. Bio/technology, 8: 939-943. |