 2000, Vol. 36
2000, Vol. 36文章信息
- 樊后保.
- Fan Houbao.
- 格氏栲群落的结构特征
- STRUCTURE FEATURES OF CASTANOPSIS KAWAKAMII COMMUNITY
- 林业科学, 2000, 36(2): 6-12.
- Scientia Silvae Sinicae, 2000, 36(2): 6-12.
-
文章历史
- 收稿日期:1998-12-07
-
作者相关文章


格氏栲(Castanopsis kawakamii Hayata)是我国三级保护植物, 在我国主要分布于福建、台湾、广东、广西、江西等地区, 多零星生长在海拔200~1000 m的常绿阔叶林中。但在福建三明小湖地区有近700 hm2以格氏栲占优势的林分, 多由百年以上的大树组成(章浩白, 1993)。为保护这一独特的森林景观, 1964年正式划界建立了三明格氏栲自然保护区。目前, 对这一罕见的森林群落的研究报道甚少(林鹏等, 1986; 郑燕明等, 1995)。因此, 有必要对格氏栲群落的结构特征及其生态原因作进一步的分析和探讨, 为科学经营和管理格氏栲种群这份珍贵的基因资源提供理论依据。
1 研究地点和方法研究地点在福建三明格氏栲自然保护区内(东经117°24′~117°27′; 北纬26°7′~26°10′)。该区位于福建武夷山东伸支脉地带, 属低山丘陵, 海拔一般在250~500 m。林下土壤主要为暗红壤, 其次为红壤和紫色土。该区属中亚热带季风型气候, 年平均气温19.5℃, 年降水量1700 mm。调查区位于小湖工区东南方的坡地上, 于1995年5、6月份, 在以格氏栲占优势的林分中, 设置25 m×40 m的典型样地10块, 分树种实测所得乔木树种(直径≥2 cm)的胸径、树高。每块样地内均匀设置5 m×5 m的样方15个,统计每样方中乔木树种的株数,详细调查灌木(包括直径<2 cm的乔木)、草本(包括蕨类植物)和层间植物的种类、株数(或多度)、高度及生长状况。在此基础上, 统计计算群落的数量指标(具体方法在文中作相应说明)。
2 结果与分析 2.1 群落种类组成 2.1.1 科属组成根据1 hm2的样地统计资料, 该群落共有维管束植物110种, 隶属于42科68属(表 1)。其中蕨类植物5种, 裸子植物1种, 双子叶植物93种, 单子叶植物11种。含单种的属占总属数的67%, 含2~3种的属占25.0%, 含4种以上的属只有栲属(4种)、榕属(4种)、樟属(4种)、石楠属(5种)和冬青属(6种)。在42个科中, 重要值在10以上的科只有壳斗科、茶科、松科、樟科、紫金牛科和冬青科, 其中壳斗科的重要值高达84.40, 占有绝对的优势。重要值在2以下的科达50%, 这说明了格氏栲群落种类组成的特点是种类丰富, 优势种显著。
|
|
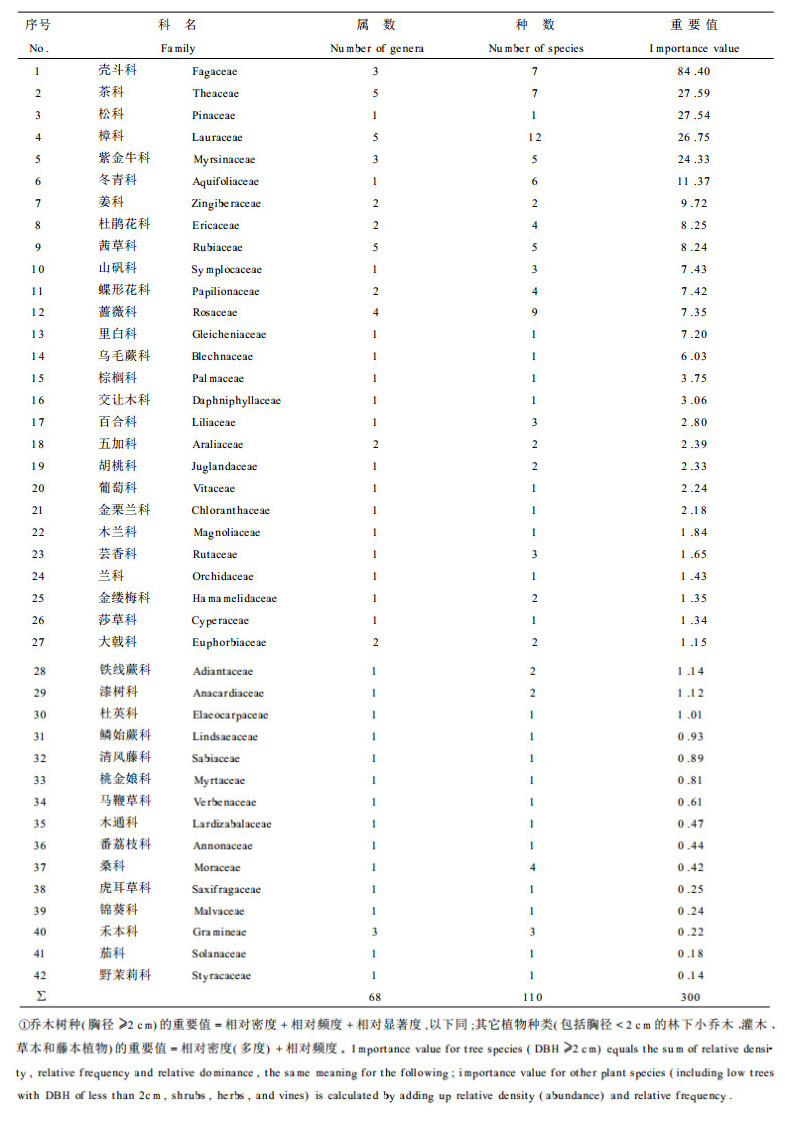
在群落的乔木树种的种类组成中, 优势种群的作用相当明显。其中格氏栲种群占有绝对的优势, 重要值高达86.08, 其相对密度、相对频度和相对显著度均居首位, 尤其是相对显著度高达61.06 (表 2)。重要值在10以上的种群只有10个。群落的基径盖度完全由上层乔木树种格氏栲、马尾松和木荷所构成, 占所有乔、灌木基径盖度的97.1%。由此可见, 该群落的森林环境主要由这3种树种所决定, 其它树种主要处于第2、第3亚层, 很少能达到20 m的高度。
|
|
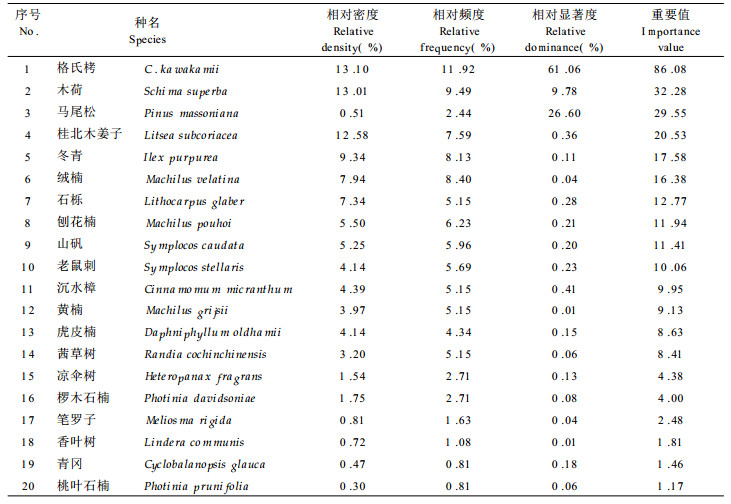
物种多样性是指种的数目及其个体分配均匀度两者的综合, 它能有效地表征生物群落和生态系统结构的复杂性(Kershaw, 1973)。本文应用Simpson指数、Shannon-Wiener指数、生态优势度和群落均匀度来描述格氏栲群落的物种多样性(表 3)。无论是Simpson指数还是ShannonWiener指数, 乔木层均最大, 灌木次之, 藤本最小。
|
|
生态优势度则是综合群落中各个种群的重要性, 反映诸种群优势状况的指标。在格氏栲群落中, 乔木层的生态优势度最小, 这说明了以种群的个体数量来衡量的话, 乔木层树种的分配比较均匀(均匀度最大)。而灌木、草本、藤本植物层的生态优势度较高, 说明了这些层次主要由少数种群所控制。如38种灌木中, 百两金(Ardisia crispa)、卡氏乌饭(Vaccinium carlesii)、狗骨柴(Tricalysia dubia) 3种群就占了总株数的52.6%。因此, 生态优势度是描述群落中各种群的作用和地位的一个综合的度量值。群落均匀度是指群落中各个种群多度的均匀程度, 它是生态优势度相反的概念。当群落生态优势度高时, 由于优势种明显, 优势种的个体数会明显多出一般种而使群落具有低的均匀度。不难理解, 群落的均匀度与物种多样性指数呈正相关关系(彭少麟, 1987; 朱守谦, 1987), 表 3也体现了这一点。
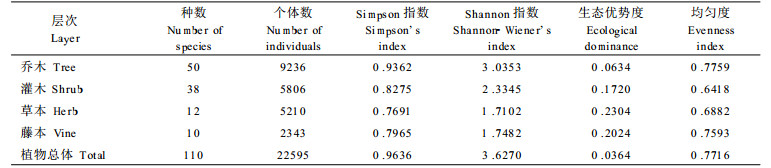
该群落植物总体的多样性指数较高, 介于南亚热带季风常绿阔叶林和中亚热带常绿阔叶林之间(表 4)。由表 4也可以看出, Shannon-Wiener's指数比Simpson指数更好地表现了群落之间的差异, 彭少麟等(1983)也指出前者更适合于亚热带常绿阔叶林。
|
|
格氏栲群落上层树种组成以格氏栲占绝对优势, 少量混生有马尾松、木荷等。树冠呈波浪状, 郁闭度约0.8, 极少出现大的林窗(forest gap)。整个群落终年浓郁苍翠, 格氏栲树干在林内的深褐色极为显目。
2.2.1 生活型根据Raunkiaer的生活型分类系统(Raunkiaer, 1934; 曲仲湘等, 1989), 该群落以包括藤本在内的高位芽植物占绝对优势, 达87.3%, 其中以中小高位芽占主导(图 1)。这一数值稍高于浙江的中亚热带常绿阔叶林(84.1%) (宋永昌等, 1982), 但明显低于云南西双版纳热带雨林(94.7%) (武吉华等,1992)。地上芽植物极少, 仅0.9%。地面芽和地下芽植物贫乏, 一年生植物缺乏。

|
图 1 格氏栲群落的生活型谱 Fig. 1 Life form spectrum of C.kawakamii community 1.大高位芽植物Megaphanerophytes; 2.中高位芽植物Mesophanerophyes; 3.小高位芽植物Microphanerophytes; 4.矮高位芽植物Nanophanerophytes; 5.藤本植物Vines; 6.地上芽植物Chamaephytes; 7.地面芽植物Hemicryptophytes; 8.地下芽植物Geophytes. |
叶特征的分析(图 2)表明, 该群落的叶级以中型叶为主, 小型叶次之, 大型叶和微型叶很少。叶型以单叶占绝对优势, 达87.3%。叶质以革质叶占优势(56.4%), 草质叶次之, 厚革质叶占有一定的比率(10.9%)。叶缘以全缘叶占多数(60%)。

|
图 2 格氏栲群落的叶特征 Fig. 2 Leaf size, leaf form, leaf texture, and leaf margin spectrums of C.kawakamii community
叶级Leaf class (a) :1 大型叶Megaphyll; 2 中型叶Mesophyll; 3 小型叶Microphyll; 4 微型叶Nanophyll. 叶级Leaf class (b) :1 单叶Single leaf; 2 复叶Compound leaf. 叶级Leaf class (c) :1 厚革质Thick leather-like; 2 革质Leatherlike; 3 草质Herbaceous; 4 薄叶Thin. 叶缘Leaf margin (d) :1 全缘Entire; 2 非全缘Non-entire. |
综上所述, 该群落的生活型和叶特征显示出一定的南亚热带季风常绿阔叶林到中亚热带常绿阔叶林的过渡地带性特点(宋永昌等, 1982; 武吉华等, 1992)。这与该地区的地理位置和气候条件有关。格氏栲自然保护区地处武夷山向东延伸的支脉末端的低山丘陵, 西北有武夷山为屏障, 削弱了冬季南侵的寒潮, 又阻截北上的海洋气流形成较丰沛的地形雨。年均气温19.4℃, 年均降水量1500 mm, 春夏季温热多雨, 冬季稍干旱, 有一定的干湿季之分, 表现出温热湿润的亚热带气候特色, 从而孕育着这一过渡地带的森林群落类型。
2.3 水平结构群落的水平结构往往决定于群落中各种群的空间格局(spatial pattern),即种群在水平空间上的配置状况(Pielou, 1988; 丁岩钦, 1980)。本文对格氏栲群落主要乔木种群(大于100株/hm2)的空间格局进行了分析(表 5)。除上层树种格氏栲种群属于随机分布(Poisson分布)外, 其它种群均属于聚集分布。其中负二项分布(negative binomial)最具代表性, 这是符合自然种群分布特点的(樊后保等, 1994; 李俊清, 1986)。应用平均拥挤度(m*)、聚块性指数(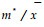
|
|
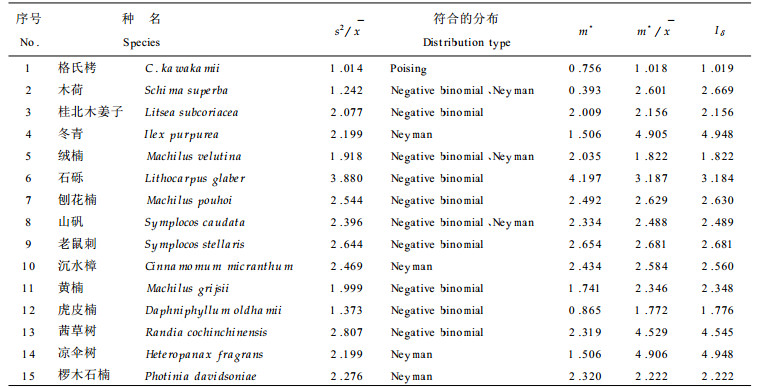
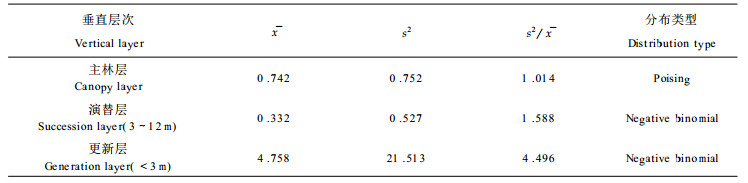
随机分布在自然界不很常见(曲仲湘等, 1989), 格氏栲主林层出现这种分布格局与其林分起源和生物学特征密切相关。三明小湖的格氏栲林的形成距今已有200多年的历史, 是由较多的格氏栲散生木天然下种在稀疏的毛竹林地上逐步形成的(章浩白, 1993)。主林层格氏栲种群大多由百年以上的大树组成, 由于长期对光照和资源竞争的结果, 只有适应性和生命力强的个体才得以保存, 形成主林层。对格氏栲种群不同垂直层次个体的空间格局作进一步分析, 可以反映出这种在时间序列上的竞争过程(表 6)。从表 6可以看出, 格氏栲种群的个体在更新层和演替层中均符合聚集分布(负二项分布), 且从更新层→演替层→主林层的过渡中, 扩散系数(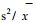
|
|
该群落的垂直层次分化比较明显, 一般可分为乔木层、灌木层和草本层, 层间植物亦比较丰富。乔木层一般可分为3个亚层, 第1亚层高约20~28 m, 种类少, 主要由格氏栲、木荷、马尾松的高大个体所组成, 形成较连续的主林层(canopy), 郁闭度0.8左右, 较少见大的林窗。其中格氏栲占绝对优势, 占总株数的72.2%, 平均树高为24.3 m, 平均胸径34.2 cm (最大一株达86.4 cm), 平均冠幅4.8 m×5.9 m。第2亚层高度约为10~16 m, 植物种类比较丰富, 主要由冬青、绒楠、石栎、刨花楠、山矾、沉水樟、虎皮楠和茜草树等组成。上层乔木树种木荷的中、幼龄个体在本亚层中占有一定的比例。本亚层树木密度大, 但基径盖度只占2.1%。第3亚层高度约为5~8 m, 由于上层乔木的幼树和下层灌木的高大植株嵌入其中, 种类组成较为复杂。主要有桂北木姜子、黄楠、梨茶(Camellia octopetala)、老鼠刺(Symplocos stellaris)等, 密度较大。
灌木层高度一般为2~4 m, 主要种类有百两金、卡氏乌饭、细齿叶柃木(Eurya nitida)、狗骨柴、毛冬青(Ilex pubescens)等, 盖度为40%~50%。在土壤湿润、林内湿度较大的中下坡位, 灌木种类较丰富, 盖度也较大。但在土壤脊薄的地方, 灌木种类较简单, 盖度小。
草本层通常分布不连续, 多集中于林缘、林窗及沟谷底。种类组成以山姜(Alpinia japonica)、狗脊(Woodwardia japonica)、芒萁(Dicranopteris dichotoma)占绝对优势。华山姜(Alpinia chinensis)、牛尾菜(Smilax riparia)、黑莎草(Gahnia tristris)等也占有一定的比例。在郁闭度较大的林分中, 草本植物种类少, 盖度也小。而在林冠破裂的空旷地或林窗中, 草本植物则比较丰富。
层间植物比较丰富, 主要由藤本植物组成, 其中以紫金牛科的网脉叶酸果藤(Embelia rudis)、酸果藤(Embelia laeta)以及蝶形花科的网络崖豆藤(Millettia reticulata)占绝对优势(总多度+频度值高达123%), 省藤(Calamus platyacanthoides)也占有较大的比例。其余有广东蛇葡萄(Ampelopsis cantoniensis)、菝葜(Smilax china)、木通(Akebia puinata)、土茯苓(Smilax glabra)、玉叶金花(Mussaenda pubescens)、南五味子(Kadsura longepedunculata)等。林内湿度不高, 除少量苔藓外未见其它附生植物。偶见乔木小板根, 但老茎生花、滴水叶尖与绞杀植物不曾见到, 雨林特征已基本消失。
3 小结三明格氏栲群落是位于中亚热带南缘的一种常绿阔叶林类型, 物种丰富, 群落结构复杂, 但优势种群的作用比较明显。优势乔木种群格氏栲控制着整个群落的森林环境, 尤以基径盖度占绝对优势。按照重要值大小, 其它优势乔木种群有木荷、马尾松、桂北木姜子、冬青、绒楠等。优势灌木种群有百两金、卡氏乌饭、细齿叶柃木、狗骨柴、毛冬青等。草本层主要由山姜、狗脊、芒萁组成。
群落的生活型谱、叶特征以及物种多样性指数的分析表明, 该群落既不同于中亚热带常绿阔叶林, 也异于南亚热带季风常绿阔叶林, 而是显示出两者之间的一种过渡类型。
上层格氏栲种群多由百年以上的老龄个体组成, 树冠已开始变稀, 林下幼树稀少, 但发现在林窗中则更新良好。有关格氏栲种群更新、演替的问题,以后再另作报道.
丁岩钦. 1980. 昆虫种群数学生态学原理与应用. 北京: 科学出版社, 84-124.
|
樊后保, 等. 1994. 蒙古栎种群空间分布格局及其动态的研究. 福建林学院学报, 14(2): 100-103. |
李俊清. 1986. 阔叶红松林中红松的分布格局及其动态. 东北林业大学学报, 14(1): 33-38. |
林鹏, 等. 1986. 福建三明瓦坑的赤枝栲林. 植物生态学与地植物学学报, 10(4): 241-253. |
彭少麟. 1987. 广东亚热带森林群落的生态优势度. 生态学报, 7(1): 36-42. |
彭少麟, 等. 1983. 鼎湖山森林群落分析Ⅰ.物种多样性. 生态科学, (1): 11-17. |
彭少麟, 等. 1989. 广东森林群落的组成结构数量特征. 植物生态学与地植物学学报, 13(1): 10-17. DOI:10.3321/j.issn:1005-264X.1989.01.011 |
曲仲湘等.植物生态学(第二版).北京: 中国教育出版社, 1989, 142~151, 158~159
|
宋永昌, 等. 1982. 浙江泰顺县乌岩岭常绿阔叶林的群落分析. 植物生态学与地植物学丛刊, 6(11): 14-34. |
王伯荪. 1987. 香港岛黄桐森林群落分析. 植物生态学与地植物学学报, 11(4): 241-250. |
王梅峒. 1988. 江西亚热带常绿阔叶林的生态学特征. 生态学报, 8(3): 249-255. |
武吉华, 等. 1992. 植物地理学(第二版). 北京: 高等教育出版社, 100-101.
|
奚为民, 等. 1993. 四川缙云山森林群落林窗边缘效应的研究. 植物生态学与地植物学学报, 17(3): 62-72. |
章浩白. 1993. 福建森林. 北京: 中国林业出版社, 107-166.
|
郑燕明, 樊后保, 等. 1995. 格氏栲种群及其主要伴生树种的空间格局. 福建林学院学报, 15(2): 97-102. |
朱守谦. 1987. 贵州部分森林群落物种多样性初步研究. 植物生态学与地植物学学报, 11(4): 286-295. |
Chapman R W. 1928. The quantitative analysis of environmen tal factors. Ecology, 9(1): 111-122. |
Kershaw K A. Quan titative and dynamic ecology.London: Edw ard Arnold, 1973, 72~142
|
Kimmins J P.Forest ecology.New York: Macmillan Publishing Company, 1987, 305~308
|
Pielou E C. (卢泽愚译).数学生态学.第二版, 北京: 科学出版社, 1988, 119~213
|
Raunkiaer C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press, 286-378.
|