 2000, Vol. 36
2000, Vol. 36文章信息
- 苏晓华, 李金花, 陈伯望, 张绮纹, 张香华.
- Su Xiaohua, Li Jinhua, Chen Bowang, Zhang Qiwen, Zhang Xianghua.
- 杨树叶片数量性状相关联标记及其图谱定位研究
- DETECTION AND IDENTIFICATION OF MOLECULAR MARKERS ASSOCIATED WITH QUANTITATIVE TRAITS OF LEAF IN POPLAR
- 林业科学, 2000, 36(1): 33-40.
- Scientia Silvae Sinicae, 2000, 36(1): 33-40.
-
文章历史
- 收稿日期:1999-01-21
-
作者相关文章


许多生理学研究已证实了叶片大小、形态、结构及着生和排列的方向性对林木生产量有重要影响(Hinckly et al., 1989; Dunlap et al., 1995), 叶片大小可以很好地预测树干材积(Leverenz et al., 1990)。但人们对林木叶片形态变异的遗传基础却知之甚少, 很难利用性状表型预测性状的遗传变异。传统的数量遗传学通常把控制数量性状的多个基因作为一个整体进行研究, 无法鉴别出单个数量性状基因以及与之有关的染色体片段更难确定数量性状基因在染色体上的位置及与其它基因的关系因而无法鉴别单个的数量性状基因位点, 并把它们定位在染色体上或采用包括基因克隆在内的现代分子生物学的方法对其进行遗传操纵(徐云碧等, 1994)。过去几十年来, 采用形态标记、细胞学标记和同工酶生化标记对QTL进行检测和作图研究, 但由于这些标记数量有限, 对生物的破坏性以及QTL分析方法的限制, 使这一领域研究长期徘徊不前。随着PCR技术的出现, 使得分子标记的获得变得容易, 极大地推动了QTL定位的理论研究。许多学者根据不同群体的遗传特性, 分别提出了标记座位与QTL相互关联的各类检测方法(惠大丰等, 1997; 李维明等, 1993)。由于RFLP和RAPD等分子标记不受数量的限制, 表型上中性, 近几年广泛应用于杨树(Bradshaw et al., 1995)、桉树(Grattapaglia et al., 1996)、火炬松(Groover et al., 1994)等的树种重要经济性状基因定位。
杨树叶片大小影响树干材积生长(Hinckly et al., 1999; Dunlap et al., 1995), 杨树杂种的叶片较大被认为是F1具有杂种优势的重要原因(Hinckly et al., 1989)。Wu等(1997)发现与杨树叶片大小有关的性状具有较大的广义遗传力, 例如树冠4个不同部位叶片性状的广义遗传力分别为:叶长宽比0.554~0.849, 叶面积0.705~0.786, 叶柄长0.812~0.869。
本研究利用数量遗传学方法和分子标记, 对美洲黑杨与青杨杂交谱系的5个叶片表型数量性状变异进行了分析, 检测出与叶片表型数量性状相互关联的RAPD标记, 并在已构建出的分子标记连锁图谱上进行了定位研究, 其目的是为实现杨树分子标记辅助选择育种提供依据及参考。
1 材料与方法 1.1 材料和分子连锁图谱采用构建分子连锁图谱的谱系三代为实验材料, 以美洲黑杨(P.deltoides Marsh.)与青杨(P.cathayana Rehd.)杂交组合产生F1, 再经F1自交产生F2群体。1996年春季在北京中国林科院苗圃种植亲本美洲黑杨(P.deltoides)和青杨(P.cathayana)、F1 (4个个体)及F2群体, 从F2群体中取80个个体用于RAPD分析及图谱的构建。苏晓华等(1998)已详细阐述了RAPD标记连锁图谱的构建, 本文不再赘述。
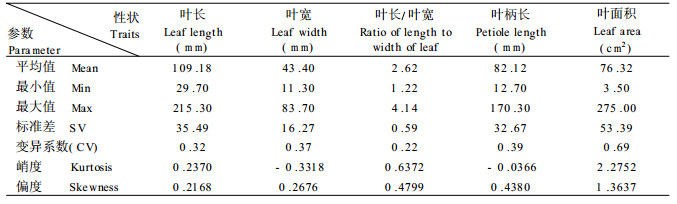
1.2 方法 1.2.1 叶片性状测定1997年秋从两根一杆平茬苗木顶端起第6片叶往下各株均取3片叶子, 测量叶长、叶宽、叶长/宽比、叶柄长和叶面积。
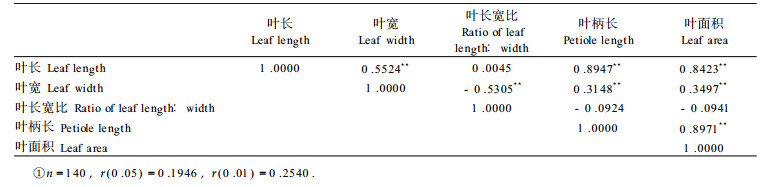
1.2.2 统计分析使用SAS统计分析软件(6.12版)对F2群体的5个数量性状进行描述统计, 得到统计参数及频率分布图, 并计算出性状间的相关系数。
1.3 数据处理根据标记电泳带型的种类将F2分离群体划分为两个亚群体:组1和组0, 应用SAS软件对180个标记位点与5个叶片数量性状表型值进行单因子方差分析, 获得与各数量性状有关的QTLs。用组间方差与总方差的比值求得这一标记产生的性状变异占性状总变异的百分率, 即标记对该性状变异的贡献率。
以用单因子方差分析检测出的与5个叶片数量性状有关的任意两个标记为因子, 根据标记两个带型的种类, 将分离群体分为4个亚群体:组01、组00、组10和组11, 对F2群体5个叶片数量性状采用双因子方差分析法求得叶片数量性状有关的两个标记互作组合。用标记互作方差与总方差的比值, 求得标记互作产生的性状变异占总变异的百分率, 即标记互作对该性状变异的贡献率。
1.4 QTLs的图谱定位在所构建的遗传连锁图谱上找出与叶片数量性状相关联的RAPD标记, 并在图谱上标出标记相关联的5个叶片数量性状。
2 结果与分析 2.1 F2代叶片数量性状表型分布与遗传变异及性状间相关系数亲本美洲黑杨叶片大, 三角型, 叶柄长; 青杨叶片小, 卵状披针形, 叶柄短。杂种F1代4个个体叶形均为中间型较一致。它们所形成的F2代叶片性状形态分化较大, 从父本型、中间型到母本型均有, 但偏父本型和偏母本型个体所占比例较小, 分别为3.25%, 9.72%, 仍以中间型居多。
F2代5个叶片数量性状的变异系数为叶长0.32, 叶宽0.37, 长/宽比0.22, 叶柄长0.39, 叶面积0.69, 性状的变异均较大(表 1)。所有叶片性状均为连续分布, 在F2群体中表现显著分离, 峭度和偏度分别为叶长0.2370, 0.2168;叶宽-0.3318, 0.26 16;叶长宽比0.6372, 0.4799;叶柄长-0.0366, 0.4380;叶面积2.2752, 1.3637;表明这5个性状的分离基本符合正态分布(图 1), 适于作遗传分析, 其中性状低值极端个体频率略高于高值极端个体频率。另外F2代还出现亲本负向遗传效应现象, 即个别单株的叶片远小于亲本青杨叶片。
|
|

|
图 1 F2代5个叶片数量性状频率分布 Fig. 1 Distribution of frequency of 5 leaf quantitative traits in F2 A叶长, B叶宽, C叶长宽比, D叶柄长, E叶面积 Leaf Length (A), leaf width (B), Ratio of leaf length:width (C), Petiole length (D), Leaf area (E). |
由F2代5个叶片数量性状间的相关系数(表 2)得知, 除叶长宽比与叶长、叶柄长和叶面积性状间不相关外, 其它性状间相关极显著, 且叶长宽比与叶宽呈负相关, 其它为正相关。
|
|
应用单因素方差分析检测出F2群体中与5个叶片数量性状相关联的标记(表 3)。
|
|

由表 3可知, 在F2群体中与叶长显著关联的标记有9个, 极显著关联的标记1个, 为RPJ07-1;这些标记座位对叶长性状变异的联合贡献率为66.23%, 其中极显著关联标记贡献率为10.54%, 占联合贡献率的15.91%。与叶宽显著关联的标记有10个, 联合贡献率为61.82%。与叶长宽比极显著关联的标记有1个, 为RPK06-2;显著关联的标记为4个; 标记联合贡献率为32.86%, 其中极显著关联标记贡献率为13.04%, 占联合贡献率的39.68%。与叶柄长极显著关联的有3个, 为RPH08-1, RPJ07-1, RPL13-1;显著关联的有6个; 标记联合贡献率为59.67%, 其中极显著关联标记贡献率为29.85%, 占联合贡献率的50.04%。与叶面积极显著关联的标记有3个, 为RPH08-1, RPL13-1, RPN05-2;显著关联的有9个; 标记联合贡献率为81.79%, 其中极显著关联标记贡献率为27.72%, 占联合贡献率的33.89% (表 3)。
2.3 与互作QTL相关联的标记应用双因素方差分析对单因素方差检测出的与叶片5个性状相关联的标记进行了任意两个不连锁的标记座位与性状的相关, 共检测出与互作QTL紧密相关联的标记座位19对(表 4)。从表 4可知, 与叶长极显著相关的有4对, 对性状的联合贡献率为40.29%;与叶宽极显著相关的有2对, 对性状的联合贡献率为17.66%;与叶长宽比极显著相关的有1对, 对性状的联合贡献率为10.84%;与叶柄长极显著相关的有5对, 对性状的联合贡献率为44.53%;与叶面积达极显著相关的有7对, 对性状的联合贡献率为82.64%。
|
|

在已构建的美洲黑杨(P.deltoides)与青杨(P.cathayana)杂种连锁图谱中, 有20个连锁群, 110个标记位点, 图谱总图距为1899.4 cM, 覆盖杨树基因组长度的70.37%, 标记平均间距为17.27 cM (苏晓华等, 1998)。与5个叶片性状相关联的标记集中分布在第4, 12, 15和17条连锁群上, 其中第2连锁群上的标记RPC06-6与叶长、叶宽、叶柄长和叶面积同时关联; 第12连锁群上的标记RPN01-2与叶长、叶宽和叶面积同时关联, 标记RPL13-1与5个叶片性状同时关联; 第15连锁群上的RPK10-4与叶长和叶柄长关联; 第17连锁群上的RPJ05-2与叶长、叶柄长和叶面积同时关联; 第15连锁群上的RPD15-3和第18连锁群上的RPK06-2与叶长和叶宽同时关联。与5个叶片性状相关联的标记个别分布在第和条连锁群上还有一些未在连锁群上(见图 2)。

|
图 2 与5个叶片数量性状相关联标记在连锁群上的位置 Fig. 2 Genetic linkage map of poplar associatel with QTL of 5 leaf quantitative traits |
大量研究证明林木叶片对林木生长量的重要性(Hinckly et al., 1989; Dunlap et al., 1995; Leverenz et al., 1990)。因此, 叶片也是林木选种的指标之一。经典数量遗传学研究表明F2代叶片数量性状间存在显著相关性, 但无法鉴别数量性状基因在染色体上的位置及其遗传效应, 也难以确定相关性状的QTL间的关系。分子标记技术的出现, 使得对林木叶片数量性状研究进入到分子水平阶段。本研究从分子遗传学角度获得证据, 为解释数量性状复杂的遗传背景和性状间的相关关系提供了很好的解释。研究发现, 与某一叶片性状相关联的标记同时与其它叶片性状关联, 与各叶片性状关联的标记位于同一连锁群或邻近的区段上, 这表明性状间存在显著相关关系的主要原因之一是存在一因多效或者是由于紧密连锁的处于同一染色体区间的不同座位控制所引起的。QTL定位的结果可以把叶片的选择分解成对叶片构成因子的选择, 在育种中利用与这些QTL连锁的标记进行辅助选择, 将有助于对林木生长量的改良。
由于RAPD和RFLP等分子标记不受数量的限制, 近几年已用于植物重要经济性状的QTL作图。目前, 在QTL作图中较常用的方法为方差分析法(analysis of variance, ANOVA)、区间作图法(interval mapping, IM)和复合区间作图法(composite interval mapping, CIM)。惠大丰等(1997)曾应用上述3种构图方法分析了玉米组合K×CML139r F2群体有关玉米螟抗性、株高和穗位高的数量性状基因(QTL)图谱, 结果3种不同方法估计的QTL位置和效应很接近, 认为ANOVA虽然难以精确确定QTL的位置, 但在发现QTL上, 仍可能是最有效的。在本研究中检测出多个QTL, 也说明方差分析法对QTL的发现能力的确很强。Wu等(1997)以毛果杨×美洲黑杨种间杂交谱系探测出叶子色素、叶柄长和叶长比率受少数QTL控制, 而叶面积、叶形、叶片着生角度、叶边缘颜色是受多数QTL控制。本项研究也检测出大量与叶面积相关联的标记, 标记数为12个, 联合贡献率为81.79%。但与叶柄长相关联的标记检测出9个, 说明叶柄长并不一定受少数QTL控制。
本研究发现, 单因子方差分析检测出的与性状显著相关联的标记, 对性状贡献率较小, 且与其它标记的互作与性状关联达极显著, 对性状的贡献率较大。这一结果的原因可能是: (1)由于特定的杂交组合F2群体所造成的, 群体特有的遗传基础决定叶片性状是由基因互作而引起的。(2)由统计方法引起的。通常使用单因子方差分析法, 只有当该QTL具有较大的单因子效应同时具有明显的互作效应时, 才能将其检测出, 而双因子方差分析可检测出有明显互作效应的QTL, 尽管该QTL的单因子效应并不显著(李春丽等, 1996)。
用于构建本研究遗传群体的杂交组合双亲在5个叶片数量性状表型上均差异较大, 但没有检测到控制它们的主基因效应, 所检测出的基因均为微效基因。这些微效基因对各自性状的联合贡献率平均在60.47%, 占性状变量的大部分, 表明该群体中可能没有主效基因。参与影响这些性状的分布基本上都只有一个主峰出现, 根据经典遗传研究方法也可推测双亲所携带的这些性状主效基因均处于等位关系, 因此该群体中在这些性状上都没有主效基因发生分离, 从而也就没能检测出它们各自的主基因效应。在其它树种QTL作图中也未检测出主效基因, 如Grattapaglia等(1996)在桉树中检测出4个树高QTLs, 联合贡献率为14%。Groover等(1994)在火炬松中检测到5个与木材比重有关的QTLs, 联合贡献率为20%。
QTL易受环境影响, QTL的效应往往与环境效应相混淆, 给检测造成了困难。另外, 当分析某一QTL时, 遗传背景中其它QTL的分离产生剩余遗传方差, 又增大了表型方差。增加重复可有效地减少样本容量、降低试验误差、提高检测QTL的灵敏度和准确性(李维明等, 1993)。本项研究正在进行增加田间重复试验研究, 以便降低环境误差, 提高QTL检测的准确性。
惠大丰, 姜长鉴, 莫惠栋. 1997. 数量性状基因图谱构建方法的比较. 作物学报, 23(2): 129-136. DOI:10.3321/j.issn:0496-3490.1997.02.001 |
李春丽, 郑康乐. 1996. 应用RAPD标记检测与水稻株高和抽穗期有关的QTLs. 遗传学报, 25(1): 34-39. |
李维明, 吴为人, 卢浩然. 1993. 检测作物数量性状基因与遗传标记连锁关系的方差分析法及其应用. 作物学报, 19(2): 97-102. DOI:10.3321/j.issn:0496-3490.1993.02.001 |
苏晓华, 张绮纹, 郑先武, 等. 1998. 美洲黑杨(Populus deltoides Marsh.) ×青杨(P. cathayana Rehd.)分子连锁图谱的构建. 林业科学, 34(6): 29-37. DOI:10.3321/j.issn:1001-7488.1998.06.004 |
徐云碧, 申宗坦, 许吉臣, 等. 1994. 利用分子标记连锁图谱进行水稻数量基因的区间作图. 中国科学(B辑), 24(9): 291-297. |
Bradshaw Jr H D, Stettler R F. 1995. Molecular genetics of growth and developmentin Populus. Ⅳ. Mapping QTLs with large effect on growth, form, and phenology traits in a foresttree. Genetics, 139: 963-973. |
Dunlap J M, Heilman P E, Stettler R F. 1995. Genetic variation in natural populations of Populus trichocarpa Torr. & Gray and its hybrids. Ⅷ Leaf and crown morphology of native black cottonwood clones from four river valleysin Washington. Canadian Journal of Forest Research, 25: 1710-1724. DOI:10.1139/x95-185 |
Grattapaglia D, Bertolucii F L G, Sederoff R. 1996. Genetic mapping of quantitative loci controlling growth and wood quality trait in Eucalyptus grandis using a maternal half-sib family and RAPD markers. Theor Appl Genet, 144: 1205-1214. |
Groover A, Devey M, Fiddler T, et al. 1994. Identification of quantitative trait loci influencing wood specific gravity in an outbred pedigree of loblolly pine. Genetics, 138: 1293-1300. |
Hinckley T M, Ceulemans R, Dunlap J M et al. Physiological, morphological, and anatomical components of hybrid vigor in Populus, In : Kreeb K. H., Richter H., Hinckley T. M., (eds), Structural and functional responses to environ mental stresses. SPB Academic PublThe Hauge, The Netherlands, 1989, 199 ~217
|
Hinckley T M, Braatne J, Ceulemans S et al. Growth dynamics and canopy structure.In : Mitchell C P, Ford-Robertson J B, Hinckley T, Sennerby-Forsse L (eds) Ecophysiology of short rotation crops. Elsevier Applied Science, London, 1992, 1 ~34
|
Larson P R, Isebrands JG. 1971. The plastochron index as applied to developmental studies of cottonwood. Can J For Res, 1: 1-11. DOI:10.1139/x71-001 |
Leverenz J W, Hinckley T M. 1990. Shoot structure, leaf area index and productivity of evergreen conifer stands. Tree Physiol, 6: 135-149. DOI:10.1093/treephys/6.2.135 |
Pereira JS, Landsberg JJ. Biomass production by fast-growing trees. NATO ASI Series. Series E: Applies science. Kluwer Academic Publ, Dordrecht, Nethersland.1989, 166 https://link.springer.com/book/10.1007/978-94-009-2348-5
|
Ridge C, Hinckley T M, Stettler R F, et al. 1986. Leaf growth characteristics of fast growing poplar hybrids Populus trichocarpa ×P.deltoides. Tree Physiol, 1: 209-216. DOI:10.1093/treephys/1.2.209 |
Wu R, Bradshqw Jr H D, Stettler R F. 1997. Molecular genetics of growth and develop mentin Populus (Salicaceae). Ⅴ. Mapping quantitative trait loci affecting leaf variation. American Journal of Botany, 84(2): 143-153. DOI:10.2307/2446076 |