 2000, Vol. 36
2000, Vol. 36文章信息
- 王华芳, 张建华, 梁建生, 尹伟伦.
- Wang Huafang, Zhang Jianhua, Liang Jiansheng, Yin Weilun.
- 侧柏和大叶相思超微弱发光土壤干旱的生理反应
- ROOT ULTRAWEAK LUMINESCENCE SUBJECTED TO SOIL DRYING IN PLATYCLADUS ORIENTALIS AND ACACIA AURICULIFORMIS
- 林业科学, 2000, 36(1): 2-8.
- Scientia Silvae Sinicae, 2000, 36(1): 2-8.
-
文章历史
- 收稿日期:1998-04-03
-
作者相关文章


2. 香港浸会大学 香港九龙塘
2. Hong Kong Baptist University Klowloong Tong, Hong Kong
植物的抗旱性已经研究了100多年, 但根系对环境干旱信息的感应与传递仍是抗旱生理的研究热点之一。土壤干旱时, 根部产生的ABA通过木质部被运送到地上器官调控其生理过程如气孔蒸腾(改变气孔导度)、光合速率和生长发育等(Davis et al., 1991)。现已认为ABA和水分是根系传递干旱信息的化学和物理信号。
最近有人提出, 植物本身存在的超微弱发光(Ultraweak luminescence, UWL)能对土壤干旱作出反应(Floryszak et al., 1992; Phyllis et al., 1993)。已有些实验支持这种观点, 如渗透胁迫的植物UWL与对照有明显区别(董家伦等, 1992), 沙生植物UWL与树种间的抗旱性有关(董家伦等, 1990)。以UWL作为光源进行光子成相, 绿豆萌发种子的胚轴生长点UWL最强(陈天鹏, 1996)。我们的工作表明侧柏根系呼吸代谢产生的对超弱发光的贡献高达这表明与植物的能量代谢相藕联, 可能是植物传递干旱信息的一种生物物理信号, 因此可以作为选择和评价树种抗旱性的一种生理指标, 但目前植物UWL对土壤干旱的生理反应还缺乏了解。本文以两个抗旱树种:侧柏(Platycladus orientalis)和大叶相思(Acacia auriculiformis)为材料, 研究其根系UWL对土壤干旱过程(急性干旱和缓慢干旱)、土壤含水量和植株含水量及水势(Ψ)等连续系统的关系, 目的是探讨UWL与土壤—植物连续系统干旱的关系, 了解UWL与土壤干旱持续时间(d)、土壤含水量和植株含水量的相关性, 为UWL能否作为选择树种抗旱性的生理指标提供实验证据。
1 材料和方法 1.1 实验材料侧柏和大叶相思为在黑色塑胶袋中土壤点播的当年生实生苗。塑胶袋直径为6.5cm, 长20cm。土壤为John Innes 2号混合基质, 其中壤土、泥炭和沙的体积比为7:3:2, 加入复合肥料15N:15P:15K6.2gL土壤-1 (该混合基质的理化性质已进行过全面测定, 是大多数实验室培植实验植物的经典配方土壤, 其水分状况通常用土壤含水量(SWC)表示即可)。每塑胶袋内播种子4~5粒, 随着种子萌发生长, 逐步间苗以淘汰长势较弱的植株, 开始进行实验处理时, 每个塑胶袋内仅剩下1株长势基本一致的健壮苗, 侧柏苗高18cm, 大叶相思苗高45cm。
1.2 实验方法 1.2.1 苗木的干旱-复水处理苗木脱去塑胶袋, 种植在土壤体积分别为3.4×10-3 m3和11.5×10-3 m3的大小明显不同的两种塑料盆里, 同时浇一次透水使土壤含水量达到饱和, 以后不再浇水使土壤自然干旱。小盆土壤干旱较大盆的慢因此形成急性干旱和缓慢干旱处理。为跟踪土壤干旱过程中的土壤含水量(SWC)以及为在植株干旱的生理状态基本相同的条件下恢复浇水, 土壤干旱处理延续到植株出现明显干旱症状如侧柏针叶卷曲略有发白, 大叶相思叶片发白下垂叶肉萎缩叶片变薄叶脉隆起时, 再恢复正常浇水。
每种植物种在小盆和大盆土壤里的植株数量不少于36株(盆)。其中18株为对照, 对照自始至终给予正常浇水并与处理植株在相同条件下培养, 培养条件为气温15~28℃, 光照强度400μmolm-2s-1, 光照时数为12h。
1.2.2 取样急性干旱和缓慢干旱处理的苗木分别每隔2d和4d取样1次, 共取样6~7和5~6次。每次取样包括:土壤、根系和木质部汁液等样品。
土壤取样按土壤学方法进行。在根系分布区(0~15cm)内, 分别取等量表土、心土和底土均匀混合以作为平均样, 每次取样为2个重复。
植物根系以水溶法溶去泥土取出, 以去离子水冲洗3次, 用剪刀剪取根尖白色致褐色部分, 以滤纸拭去根系表面的残留土壤, 立即称取200mg装入LS6500液体闪烁计数器(Liquid scintillation counting system, LSC)专用的闪烁瓶中, 在室温(23℃)下避光4.5h方可测定其超弱发光(UWL)。
1.2.3 根系超弱发光(UWL)的测定根系超弱发光可以用生物发光仪也可以用LSC测定, 本次实验以Beckman LS6500 LSC单光子监测器(Single photon monitor)测定, 按照该仪器说明书设置仪器参数如下:
计算模型:单光子检测器; 计数时间/样品:1.00;数据采集数目:20;计数时间/数据点:0.05;计数样品设置:1;补偿因子:1.000000。
被测样品的数量、处理方法、预处理时间和数据的解释单位等是在测定样品之前进行预备实验来确定(王华芳等, 1994)。
根系样品在测定UWL之后, 置于105℃烘箱内烘致恒重并称量。测定数据扣除本底求出单位重量的UWL强度, 即: (样品cpm-测样瓶cpm) /样品干重(cpm为每分钟计数), 再求出重复样品(2个重复)超弱发光平均强度, 以对照植株UWL强度为100%进行数据归一化。
1.2.4 植株水势(Ψ)测定植株水势用压力室法测定, 将植株从离表土5cm高处剪断, 立即将地上部分装入预先被水汽饱和的塑料袋中, 使切口部分的茎伸出塑料袋3cm, 将塑料袋包裹整个枝条并置于压力室内以压缩N2加压, 仔细观察枝条切口表面, 以初见其返潮时的气体压力为植株水势(MPa)。压力室为Model 300, Soil Moisture Equipment Co.USA。塑料袋内的饱和水汽是在测定植株水势之前, 往袋内放一片湿的滤纸保持5~10min即可达到饱和。
1.2.5 土壤含水量(SWC)测定SWC测定按土壤学的常规方法进行, 土壤样品称鲜重之后, 置于105℃烘箱内烘致恒重并称量。按以下公式计算SWC:
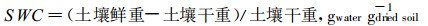
|
侧柏和大叶相思根系UWL随土壤干旱持续时间(d)的动态见图 1。根系UWL与土壤干旱持续时间显著负相关(P < 0.001)。随着土壤干旱持续时间延长, 侧柏根系UWL从1.4×106±1.4×104 cpm gdried weight-1 (100%)呈对数式衰减, 小盆和大盆土壤分别干旱19d和24d, 根系UWL分别下降为20%和30%;大叶相思根系UWL随土壤干旱持续时间(d)从4×106±7.9×104 cpmgdried weight-1 (100%)呈直线式衰减。小盆和大盆土壤分别干旱11d和16d, UWL衰减为31%。缓慢干旱比急性干旱的同等情况晚6d左右。土壤缓慢干旱过程有利于在较长时间内维持侧柏和大叶相思根系较高的UWL水平。

|
图 1 侧柏(△ ▲)和大叶相思(● ○)根系UWL对土壤急性干旱(——)和缓慢干旱(……)干旱持续时间的反应箭头之后为复水的情况 Fig. 1 Changes of ultraweak luminescence of P.orientalis
(△ ▲)and A.auriculiformis(● ○)subjected to either rapid (——)or slow (……) soil drying.Rewatering was supplied and indicated by arrows.Values mean per plant.
▲ y =262 / [ 1 + (x/529) 0.2] -162, n =17, R2 =0.9446 △ y =346 / [ 1 + (x/529) 0.3] -232, n =18, R2 =0.9703 ● y =99-6 x, n =18, R2 =0.9644 ○ y =103-5 x, n =15, R2 =0.9458 |
侧柏和大叶相思根系UWL与土壤含水量(SWC)显著相关(P < 0.001), 见图 2。随着SWC的减少, 侧柏根系UWL呈指数式衰减。小盆土壤SWC从0.24gwatergdried soil-1下降63%, UWL急剧衰减70%, 之后则随着SWC减少缓慢衰减; 大盆土壤SWC减少58%, UWL衰减50%。大叶相思小盆土壤根系UWL随着SWC下降呈对数式衰减。SWC开始减少9%, UWL缓慢衰减20%, 之后SWC减少20%, UWL迅速衰减50%;大盆土壤的根系UWL随SWC减少呈线性衰减, SWC减少58%, UWL衰减70%。土壤缓慢干旱的侧柏根系比急性干旱的保持较高的UWL水平, 但大叶相思根系UWL衰减急性干旱的快, 其原因还需要进一步研究。

|
图 2 侧柏(▲ △)和大叶相思(● ○)根系UWL与土壤急性干旱(——)和缓慢干旱(……)的含水量(SWC)的关系 Fig. 2 Relationship between root ultraweak luminescence and soil water co ntent (SWC) in P.orientalis (▲ △) and A.auriculi formis(● ○) subjected to either rapid (——)or slow (……) soil drying, Values mean per plant.
▲ y =5.4Exp11.57 x, n =26, R2 =0.7739 △ y =100Exp6.6 x, n =24, R2 =0.8982 ● y =215 x/ (0.2 +x) -23, n =24, R2 =0.8504 ○ y =281 x -53, n =21, R2 =0.8753 |
侧柏根系UWL与根系含水量(RWC)显著相关, P < 0.001, 见图 3。在正常SWC条件下侧柏根系RWC为其干重的70%。在土壤干旱条件下, 根系UWL都随着SWC降低呈指数式衰减, 但随RWC降低, 土壤缓慢干旱的侧柏根系UWL衰减较急性干旱的慢。

|
图 3 侧柏根系UWL对急性干旱(▲——)和缓慢干旱(△……)的根系含水量(RWC)的反应 Fig. 3 Relatio nship between root ultraweak luminescence and root water content(RWC) in P.orientalis subjected to either rapid (▲——) or slow(△……) soil drying.Values mean per plant. ▲ y =17Exp(2.5 x), n =12, R2 =0.9645 △ y =28Exp(1.7 x), n =12, R2=0.9523 |
侧柏和大叶相思根系UWL与植株水势显著相关, P < 0.1。植株水势(Ψ)随土壤干旱时间的动态见图 4。在正常情况下Ψ为-1MPa以上, 随着土壤干旱进程Ψ在对数式下降, 而随着土壤复水进程以对数式递增。植株根系UWL随Ψ下降呈指数式衰减, P < 0.1, 见图 5。侧柏小盆和大盆植株分别干旱19d和24d, Ψ从-1MPa分别降到-4MPa和-2.5MPa, 根系UWL分别递减90%和70%。大叶相思小盆和大盆土壤分别干旱11d和16d, Ψ分别下降到-3.2MPa和-2MPa, UWL分别衰减70%和80%。缓慢干旱的大叶相思根系UWL随Ψ衰减较快其原因仍需要研究。

|
图 4 侧柏(▲ △)和大叶相思(● ○)植株水势(Ψ)随土壤急性干旱(——)和缓慢干旱(……)时间的动态, 箭头之后表示复水的情况。 Fig. 4 Changes of leaf water potential(Ψ) of P.orientalis (▲ △)and A.auriculiformis (● ○)subjected to rapid (——)or slow(……) soil drying.Rewatering was supplied and indicated by arrows.Values mean per plant. |

|
图 5 侧柏(▲ △)和大叶相思(● ○)根系UWL与土壤急性干旱(——)和缓慢干旱(……)时植株水势(Ψ)的关系 Fig. 5 Relationship between root ultraweak luminescence and leaf water potential(MPa) in P.orientalis(▲ △) and A.auriculiformis(● ○) subjected to rapid(——) and slow(……) soil drying. Values mean per plant.
▲ y = 147Exp x, n = 26, R2 = 0.37.5 △ y = 191Exp0.9 x, n =19, R2 =0.468 ● y =114Exp0.5 x, n =18, R2 =0.8394 ○ y = 120Exp0.8 x, n =15, R2 =0.7967 |
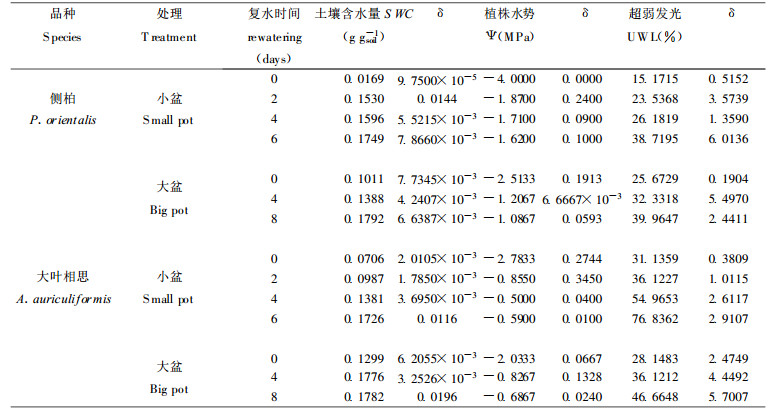
侧柏和大叶相思根系UWL对干旱土壤复水有反应, 见表 1。干旱土壤复水, SWC逐渐得到恢复土壤复水6~8d, 已恢复到干旱处理之前的75%左右, 植株水势也接近正常水平但根系UWL的恢复到40%, 特别是缓慢干旱的侧柏根系UWL在土壤复水的前4d回升1%, 其后的恢复较迅速, 到第8已回升到36%。从总的情况看, 植株根系UWL对SWC和Ψ的恢复有协同作用, 但UWL的恢复后于SWC和Ψ。
|
|
以上结果表明, 侧柏和大叶相思根系超弱发光(UWL)强度与土壤干旱持续时间和土壤含水量显著相关(图 1~3和图 5), 根系UWL对土壤干旱的反应非常灵敏。当植株受到土壤干旱胁迫时, 根系UWL和SWC均随土壤干旱持续时间下降, 但根系UWL衰减速率比SWC快; 解除土壤干旱胁迫时, 根系UWL随SWC恢复而增强。这表明, 植株根系UWL能对土壤干旱信息作出迅速反应, 可以作为评价树种抗旱性的指标引入林业。UWL与植株生理活动相藕联, 对土壤水分变化的反应通过生物化学分子的能级跃迁(Popp. et al., 1992), 以UWL光子的形式反映出来(Phyllis et al., 1993)。因此, 以其作为植株抗旱性的综合指标有其生物学基础, 具有很高的可靠性。UWL的测定是以光电倍增管为基础, 与测定土壤含水量的称重法或测定水势的压力室方法相比, 其灵敏度非常高。
侧柏和大叶相思根系UWL与RWC相关(P < 0.001, 图 3), 水分以被动吸收过程从土壤进入根系, 土壤水分很容易与根系无阻空间的水分达到动态平衡, 形成土壤-根系水分连续系统。因此, 根细胞对土壤水分的感应首先是通过无阻空间传导, 其UWL与SWC显著相关也可以说与根系无阻空间的水分显著相关。
根系UWL对SWC反应灵敏而对Ψ的反应较不敏感。分析其理由主要有以下3方面: (1)比较SWC与根系UWL和Ψ的关系可知, 在土壤干旱条件下随着SWC下降, 根系UWL呈指数式递减(见图 6), SWC从正常水平下降时微小的变化即可引起UWL较明显的反应; Ψ随SWC呈对数式变化, 见图 7, SWC从正常水平下降时变化较平稳, 微小的变化不足以引起Ψ的明显响应, 只有SWC无限小时Ψ的变化才显著即对SWC反应灵敏。(2)根系UWL的产生与根系的呼吸代谢相藕联, 氧化磷酸化过程产生的ATP对UWL的贡献高达80% (毛大璋等, 1988), 土壤干旱直接导致根系的呼吸速率下降, 氧化磷酸化解藕联, ATP生成减少以至UWL衰减, 该过程与Ψ没有直接联系; (3)尽管植株水势是反映其水分状况的重要指标, 但水势的变化与植株体内的水分总量有关, 其变化受体内水分总量的缓冲因此相当缓慢; 另一方面, 目前测定水势的较好方法是压力室法, 其精度和灵敏度较低, 难以与测定UWL的光电倍增管的灵敏度相比。UWL对土壤干旱的敏感性应该从植株生理生化方面进一步探讨(Popp, 1988)。

|
图 6 侧柏(▲ △)和大叶相思(● ○)植株根系UWL随土壤急性干旱(——)和缓慢干旱(……)的含水量(SWC)变化的总趋势 Fig. 6 General relationship between root ultraweak lumines cence and soil water content(SWC) in P.orientalis(▲ △) and A.auriculiformis(● ○) subjected to either rapid(——) or slow(……) soil drying Values mean per plant. y =16Exp7.2 x, n =71, R2 =0.6793 |

|
图 7 侧柏(▲ △)和大叶相思(● ○)植株水势随土壤急性干旱(——)和缓慢干旱(……)的含水量(S WC)变化的总趋势 Fig. 7 General relationship betw een leaf water potential and soil water content(SWC) in P.orientalis(▲ △)and A.auriculiformis(● ○) subjected to either rapid(——)or slow (……) soil drying Values mean per plant. y =5.8 x/(0.1 +x)-4.7, n =71, R2=0.7547 |
根系UWL的绝对强度不能反应侧柏和大叶相思的抗旱性。但是可以比较其相同条件下的相对强度来判断树种抗性。这和植株呼吸速率等生理生化指标反应植株抗性的情况一样, 因此UWL反应树种抗性的结果与用其他方法测定的结果一致。但UWL作为生理生化反应的综合指标, 其测定方法简单方便灵敏度高(本次实验可以测到105cpm/gdried weight), 不仅可以用来反应植株的一般抗旱性, 还可以用来研究其他方法难以反应或不能反应的干旱胁迫早期的植株抗旱性。但是UWL作为高灵敏的树种抗性指标用于实际还望有更多的研究数据和资料的支持.
陈天明, 俞信, 王苏生. 1996. 超微弱生物发光图象中的统计检验. 光学学报, 16(6): 806-811. DOI:10.3321/j.issn:0253-2239.1996.06.021 |
董家伦, 李树真. 1992. 液闪与超弱发光农用技术. 兰州: 甘肃科学技术出版社.
|
董家伦, 李树真. 1990. 一些沙生植物苗期超弱发光特征研究. 中国沙漠, 10(2): 29-33. |
毛大璋, 沈恂, 张月敬, 等. 1988. 代谢抑制剂对萌发绿豆超弱发光的影响. 生物物理学报, 4(2): 116-120. |
杨启简. 1987. 大豆、小麦、玉米的UWL与干旱条件下种子萌发的关系. 生物化学与生物物理进展, 16(6): 452-454. |
王华芳, 路永斌, 卫蓉. 1994. 树木辉光的测定. 北京林业大学学报, 16(增刊2): 41-44. |
Davis WJ, Zhang J. 1991. Root signals and the regulation of growth and development of plants in drying soil. Annu Rev Plant Physiol Plant Mol Biol, 42: 55-76. DOI:10.1146/annurev.pp.42.060191.000415 |
Floryszak W J, Grabikowski E, Kubis J, et al. 1992. The effect of spermidine on lipid peroxidation in wheat leaves during water stress. Acta Physiologiae Plantarum, 14(1): 3-10. |
Phyllis LD, Szalay AA. 1993. Use of bioluminescence to monitor osmotic response in transformed Nictiana tabacum suspension cell tures. J Biolumi Chemilumi, 8: 77. |
Popp FA. 1988. Biophoton emission. Experientia, 44(7): 543-544. DOI:10.1007/BF01953300 |
Popp FA, Li KH, Gu Q. 1992. Recent advancesin biophoton research and its applications. World Scientific Publishing Co. Pte. Ltd..
|
White MR, Craig FF, Watmore D, et al. Applications of the direct imaging of firefly luciferase expression in single intact mam milla cell using charge-coupled device cameras. In Stanly PS, Krica LJ. Bioluminescence and chemiluminescence current status. John Wiley & Sons, 1991 : 357 ~364
|