 2009, Vol. 45
2009, Vol. 45文章信息
- 何飞, 刘兴良, 马钦彦, 史作民, 陈俊华.
- He Fei, Liu Xingliang, Ma Qinyan, Shi Zuomin, Chen Junhua
- 横断山区野桂花林群落特征
- Community Characteristics of Osmanthus yunnanensis Forest Distribu ting in Hengduanshan Mountain
- 林业科学, 2009, 45(6): 153-157.
- Scientia Silvae Sinicae, 2009, 45(6): 153-157.
-
文章历史
- 收稿日期:2007-10-11
-
作者相关文章



2. 四川省林业科学研究院 成都 610081;
3. 中国林业科学研究院森林生态环境与保护研究所 国家林业局森林生态环境重点实验室 北京 100091
2. Sichuan Academy of Forestry Chengdu 610081;
3. Institute of Forest Ecology, Environment and Protection, Chinese Academy of ForestryKey Laboratory on Forest Ecology and Environment of State Forestry Administration Beijing 100091
野桂花(Osmanthus yunnanensis)属木犀科(Oleaceae),特产于横断山区的四川西部、云南西北部和西藏的东南部(张美珍等,1992),在四川分布的范围最广,分布于海拔1 350~2 800 m的山坡或沟边,常混生于森林中,很少形成群落。目前只发现野桂花在四川省的宝兴县东拉山、泸定县海螺沟、大邑县的西岭雪山以及崇州县的鸡冠山等地有群落分布,其中宝兴县东拉山的野桂花林的分布面积最大。世界木犀属植物共有31个种,80%分布在东亚地区,中国有26个种,是该属的分布中心(向其柏等,2004)。中国木犀属植物中,仅野桂花和普通桂花(Osmanthus fragrans)有天然群落分布。据报道,普通桂花天然群落分布于福建省长汀县(董建文等,2002)、湖南省浏阳市(易继君,2006)、江西省庐山(汪德娥等,1997)和浙江省淳安县(王贤荣等,2007)等地,野桂花仅在横断山区的四川西部形成群落。关于普通桂花天然群落的研究很少,主要有种群生命表及生存分析(林勇明等,2007b)、群落物种数量特征研究(董建文等,2002;王贤荣等,2004)、群落主要树种种间关联及其对混交度的响应研究(林勇明等,2007a)以及群落特征的初步研究(汪德娥等,1997;郝日明等,2005;易继君,2006;王贤荣等,2007), 对野桂花群落特征的研究更少。
野桂花和普通桂花有明显不同:1)野桂花在4—5月开花,花黄白色,芳香,而普通桂花多数品系8月开花;2)野桂花适合于较高海拔和较高纬度的城乡生长,分布于海拔2 000 m处的较湿润地区,而普通桂花低海拔分布且不适宜北方城市生长。为深入认识野桂花的群落特性,丰富我国桂花植物的研究资料,为该种的引种驯化和资源开发提供理论依据,有必要对分布于横断山区宝兴县东拉山的野桂花林群落特征进行研究。
1 研究区概况研究区位于四川省横断山区东拉山,属宝兴县陇东镇(102°31′E, 30°22′N),距宝兴县城约46 km,为高山峡谷地貌类型。距研究地15 km处的硗碛乡泥巴沟气象站观测(海拔2 600 m)资料(2006年)表明,该区年降水量1 304.7 mm,年平均气温7.2 ℃,1月平均气温-3 ℃,7月平均气温16.5 ℃,年均相对湿度79%,全年日照时数1 921 h,全年无霜期296 d。研究区主要土壤类型为山地黄壤和黄棕壤。在中国植物区系划分中,该区域属于泛北极植物区,中国-喜马拉雅森林植物亚区中的横断山脉植物地区(吴征镒,1979)。植被类型有常绿阔叶林、常绿和落叶阔叶混交林、落叶阔叶林、针阔混交林等。野桂花呈小团块状分布于该区海拔1 100~2 300 m范围内的沟谷和山坡,分布面积近2 000 hm2。该树种最大胸径100.64 cm,高度可达30 m,历年花盛期为4月中旬,花期能持续1个月左右,8月中下旬果实成熟。
2 研究方法采用样方法进行群落调查(林鹏,1986;何飞等,2001;史作民等,2005):在初步踏察的基础上,兼顾群落分布的海拔和坡向,选择典型性地段共设12个15 m×10 m的长方形样地,记录每个样地的坡向、坡位、海拔和土壤类型等,样地概况见表 1。在每个样地内,对胸径≥3 cm的乔木进行每木检尺,识别出乔木种名,并测其树高、枝下高、冠幅和胸径;在每个样地内按对角线法设置5个2 m×2 m的灌木样方和5个1 m×1 m的草本样方:在每个灌木样方内识别出种名,测定每种的高度、冠幅和基径;草本样方同样识别出种名,测出每种的高度和盖度。
|
|
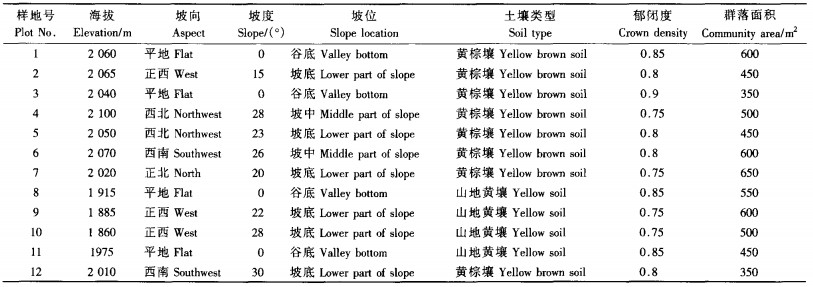
群落生活型采用Raunkiaer(1907)生活型分类系统(林鹏,1986);叶型按Raunkiaer(1934)的叶型分类系统(王伯荪,1987);重要值=(相对多度+相对频度+相对显著度)/300(王伯荪,1987)。
3 结果与分析 3.1 群落季相变化野桂花群落镶嵌在常绿和落叶阔叶混交林、落叶阔叶林或针阔混交林中,乔木层以常绿的野桂花为优势种,上层伴生植物主要有槭树(Acer spp.)、三桠乌药(Lindera obtusiloba)、西南樱桃(Prunus pilosiuscula)和领春木(Euptelea pleiospermum)等落叶植物,季相变化非常明显。春季时,在小块状“绿岛”的野桂花群落里有春季开花的杜鹃(Rhododendron spp.)、樱桃(Prunus spp.)及木兰(Magnolia spp.)加上淡红和黄绿色的嫩叶,色彩斑驳;夏季植物色泽浓绿,树冠连接,景观呈现深绿色,时有或红或白的杜鹃(Rhododendron spp.)(夏天开花)开放,群落生机盎然;秋季落叶树开始落叶,红、黄相间的彩叶点缀于“绿岛”上部;冬季呈“岛”状分布的野桂花群落仍以绿色为主景渡过寒冬。
3.2 群落生活型组成根据Raunkiaer(1907)生活型分类系统,结合本群落特点进行统计分析。该群落高位芽植物占绝对优势,有87种,占总数的54.04%,其中大高位芽植物1种,中高位芽植物27种,小高位芽植物29种,矮高位芽植物30种,它们是构建本群落的主体;其次是地下芽植物占相对较高的比例,有43种,为总数的26.71%,在地下芽植物中,蕨类植物就有17种,说明群落生境湿润;然而,地面芽植物和地上芽植物各为17和8种,各占10.56%和4.97%;一年生种子植物只有6种,占3.73%。此外,藤本植物和附生植物也占有相对较高的比例,反映出群落生境比较湿润。在藤本植物中,有巨型藤本植物存在,如钻地风(Schizophragma integrifolium)、狗枣猕猴桃(Actinidia kolomikta)、五叶瓜藤(Holboellia fargesii)和阔叶清风藤(Sabia latifolia)等,显示出群落的古老性。总体来看,野桂花林群落生活型分布格局反映了该地具有阴冷、湿润的气候特点,有利于中高位芽、小高位芽、矮高位芽、地下芽和地面芽植物的生长与分化。
3.3 群落植物叶的性质按Raunkiaer的叶型分类系统来进行叶型分析,该系统根据叶面积大小将植物叶分为6级(王伯荪,1987),各级间相差9倍。1级:微叶,叶面积0~25 mm2;2级:细叶,叶面积26~225 mm2;3级:小叶,叶面积226~2 025 mm2;4级:中叶,叶面积2 026~18 225 mm2;5级:大叶,叶面积18 226~164 025 mm2;6级:巨叶,叶面积>164 025 mm2。如表 2所示,野桂花群落以4级中型叶为主,共计80种,占49.69%;其次是3级小型叶和5级大型叶,分别为38和29种,各占总数的23.60%和18.01%。中、小型叶占主要部分,基本上反映了群落位于山地常绿阔叶林与高山暗针林之间的植被带的叶级谱性质。群落大型叶以蕨类植物为主,而且十分丰富,有13种,显示了野桂花群落生境的湿润性。
|
|

野桂花群落以单叶为主,共计120种,占74.53%;其余为复叶,共有41种,占25.47%。叶质中草质占优势,有107种,占66.46%;革质有54种,占33.54%。群落以草质叶为主,反映出该群落大多数伴生植物种具有山地温带落叶的特点;革质叶占有一定比例,反映出群落含有低海拔的常绿成分。叶缘为全缘的有60种,占37.27%;非全缘为101种,占62.73%(见表 3);2类叶缘兼有的有6种:三桠乌药、常春藤(Hedera nepalensis var.sinensis)、水红木(Viburnum cylindricum)、野桂花、三褶脉紫菀(Aster ageratoides)和钻地风。
|
|

野桂花群落结构简单、层次分明,可分为乔木层、灌木层、草本层和层间层。乔木层高3~18 m,可分3个亚层。乔木Ⅰ亚层高13~18 m,主要树种有巴山榧(Torreya fargesii)、铁杉(Tsuga chinensis)、中华槭(Acer sinense)、三桠乌药、西南樱桃、领春木(Euptelea pleiospermum)、疏花槭(Acer laxiflorum)、凹叶木兰(Magnolia sargentiana)和野桂花9种,此层共有62株乔木,盖度30%~35%,野桂花有10株,最高14.5 m,最大胸径22.3 cm,此层主要为落叶树种和针叶树种。乔木Ⅱ亚层高8~13 m,盖度60%,种类除少数Ⅰ亚层树种外,以野桂花为主,阴性树种有所增加,如宝兴梾木(Cornus scabrida)和红果树(Stranvaesia davidiana)等,此层组成树种12种共234株,野桂花168株,占此层总株数的71.79%,野桂花平均树高11.8 m,平均胸径12.73 cm。乔木Ⅲ亚层高3~8 m,共有13种172株,野桂花114株,占此层总株数的66.28%,伴生有猫儿刺(Ilex pernyi)、小蜡树(Ligustrum sinense)、长叶溲疏(Deutzia longifolia)、卵叶钓樟(Lindera limprichtii)、垂枝泡花树(Meliosma flexuosa)和喜丛柳(Salix driophila)等耐荫树种。灌木层在乔木层覆盖下,阴暗潮湿,多数种类呈丛状分布,盖度15%~20%,野桂花幼苗零星分布在母树周围,据小样方(2 m ×2 m)调查推算每公顷幼苗达900余株,耐荫灌木种有鞘柄菝葜(Smilax stans)、八宝茶卫矛(Euonymus przewalskii)、小叶忍冬(Lonicera microphylla)、云南冬青(Ilex yunnanensis)、杜鹃(Rhododendron spp.)、悬钩子(Rubus spp.)、青荚叶(Helwingia japonica)和铁籽(Myrsine africana)等。草本层植物丰富,共有46种,盖度10%~15%,以蕨类植物为主,反映了群落生境湿润。层间层植物极其丰富,共有18种,如常春藤、粉背南蛇藤(Celastrus hypoleucus)、金刚藤(Berchemia floribunda)、钻地风、淮通马兜铃(Aristolochia moupinensis)、华中五味子、巴东忍冬(Lonicera henryi)等,有些种类在该群落中形体巨大,显示出野桂花林的古老性。
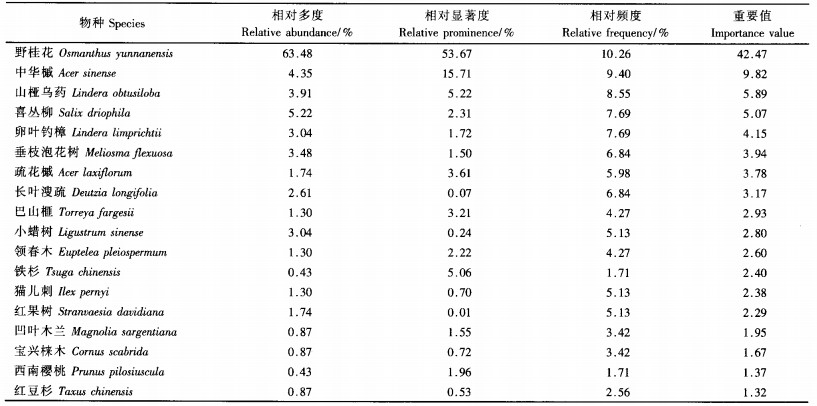
3.5 树种优势度12个150 m2的样地资料统计结果表明,组成野桂花群落的乔木(DBH≥3 cm)共计18种,隶属于13科。通过重要值排序得到表 4,由表 4可知,野桂花的优势度很显著,它的相对多度(63.48%)、相对显著度(53.67%)和重要值(42.47)都远远大于其他树种,是乔木层的优势种。其他树种如疏花槭、铁杉、凹叶木兰、三桠乌药等,虽然形体高大,但株数甚少,优势度不显著,是群落的伴生种。
|
|
种群数量及其空间分布的变动是群落动态的重要标志,而年龄结构又是种群变动的重要特征。由于实际调查中很难获得树木个体的确切年龄,本研究采用株数与胸径和树高的相互关系分析野桂花种群的动态变化规律。在12个样地中,胸径≥3 cm且树高≥4 m的野桂花共有292株。从图 1树高结构图可知,树高在6 m以下的有56株,占野桂花总株数的19.18%;树高在8 m以下的有114株,占野桂花总株数的39.04%;树高增加到10 m时,株数急升到192株,占野桂花总株数的65.75%,在此树高级上株数最多;此后随着树高增加,野桂花株数减少。从图 1胸径结构图可知,胸径在6 cm以下的野桂花有64株,占野桂花总株数的21.92%;胸径在12 cm以下的有192株,占野桂花总株数的65.75%;之后,随着胸径的增加,各胸径级的株数也逐渐减少。从图 1可见,野桂花种群年龄结构基本上呈现正态分布规律,即由多数中龄个体和少数老、幼龄个体组成相对稳定的种群结构,显示出该群落处于相对稳定的演替阶段。

|
图 1 树高和胸径分布 Figure 1 Distribution of tree height and DBH |
野桂花群落是横断山区四川西部特有的植物群落,该群落季相变化非常明显,以高位芽植物占绝对优势,群落生境湿润,具有耐冬季酷寒气候的生活习性,地下芽植物占相对较高的比例,蕨类植物很丰富,地面芽植物也占有很大比例。群落中植物叶的性质以中型叶、单叶、非全缘叶、草质叶为主,群落以草质叶为主,反映出该群落大多数伴生植物种具有山地温带落叶的特点;革质叶占有一定比例,反映出群落含有低海拔的常绿成分。野桂花群落乔木层中,野桂花的重要值最大,是群落的优势种。该群落结构简单、层次分明,可分为乔木层、灌木层、草本层和层间层,在群落垂直结构的各个层中,野桂花在乔木Ⅰ,Ⅱ和Ⅲ亚层中分别有10,168和114株,在灌木层和草本层中也有大量的幼树和幼苗,表明野桂花种群处于相对稳定的演替阶段。此外,野桂花种群年龄结构基本上呈现正态分布规律,进一步论证了野桂花种群演替正处于相对稳定阶段。
木犀属树种多在秋季开花,(刘玉莲,1993)而木犀属的野桂花是特产于横断山区的中海拔地区的常绿树种,春夏之交开花,具有一定耐寒习性,适合作为一些较高海拔和较高纬度的城乡绿化树种。鉴于野桂花具有重大的科研和生产价值,应当对其扦插、嫁接、种子育苗、组织培养、杂交育种、物候期、遗传多样性、引种驯化、适生区域和生长环境等方面进行研究。
董建文, 范小明, 吴东来, 等. 2002. 福建长汀石峰寨景区桂花次生林群落物种数量特征. 植物资源与环境学报, 11(4): 40-44. DOI:10.3969/j.issn.1674-7895.2002.04.009 |
郝日明, 臧德奎, 向其柏. 2005. 湖南省浏阳市周洛村桂花峡野生桂花资源调查. 园艺学报, 32(5): 926-929. DOI:10.3321/j.issn:0513-353X.2005.05.038 |
何飞, 郑庆衍, 刘克旺. 2001. 江西宜丰县官山穗花杉群落特征初步研究. 中南林学院学报, 21(1): 74-77. DOI:10.3969/j.issn.1673-923X.2001.01.019 |
林鹏. 1986. 植物群落学. 上海: 上海科学技术出版社, 48-90.
|
林勇明, 洪滔, 吴承祯, 等. 2007a. 桂花次生群落主要树种种间关联及其对混交度的响应. 应用与环境生物学报, 13(3): 327-332. |
林勇明, 洪滔, 吴承祯, 等. 2007b. 桂花野生种群生命表及生存分析. 北京林业大学学报, 29(3): 185-188. |
刘玉莲. 1993. 桂花品种分类及木犀属种质资源的利用. 植物资源与环境, 2(2): 44-48. |
史作民, 程瑞梅, 刘世荣, 等. 2005. 河南宝天曼化香林特征及物种多样性. 山地学报, 23(3): 374-380. DOI:10.3969/j.issn.1008-2786.2005.03.019 |
汪德娥, 王宗海. 1997. 庐山山南桂花结实习性及野生群落研究. 林业科技通讯, 10: 18-21. |
王伯荪. 1987. 植物群落学. 北京: 高等教育出版社, 25-65.
|
王贤荣, 陈昕, 季春峰. 2007. 千岛湖桂花岛桂花群落特征初步研究. 林业科技开发, 21(5): 22-24. DOI:10.3969/j.issn.1000-8101.2007.05.007 |
王贤荣, 袁发银, 陈昕, 等. 2004. 湖南浏阳桂花峡桂花次生林的物种数量特征. 南京林业大学学报:自然科学版, 28(增刊): 41-44. |
吴征镒. 1979. 论中国植物区系的分区问题. 云南植物研究, 1(1): 1-22. |
向其柏, 刘玉莲. 2004. 桂花资源的开发与应用现状及发展趋势. 南京林业大学学报:自然科学版, 28(增刊): 104-108. |
易继君. 2006. 浏阳周洛天然桂花林群落特征. 湖南林业科技, 33(3): 68-69. DOI:10.3969/j.issn.1003-5710.2006.03.024 |
张美珍, 邱莲卿. 1992. 中国植物志:第六十一卷. 北京: 科学出版社, 101-103.
|