 2009, Vol. 45
2009, Vol. 45文章信息
- 赵颖, 周志春, 金国庆.
- Zhao Ying, Zhou Zhichun, Jin Guoqing
- 马尾松苗木生长和根系性状的GCA/SCA及磷素环境影响
- GCA/SCA of Seedling Growth and Root Parameters in Pinus massoniana and the Phosphorus Environment Influence
- 林业科学, 2009, 45(6): 27-33.
- Scientia Silvae Sinicae, 2009, 45(6): 27-33.
-
文章历史
- 收稿日期:2008-05-09
-
作者相关文章



磷是植物生长发育不可缺少的一种大量营养元素。我国土壤虽具有巨大的磷库,但其有效磷含量却很低,如华南地区低磷酸性红壤的有效磷含量仅为2 mg·kg-1(李庆逵,1985)。在生产上虽可通过施用磷肥提高土壤有效磷含量,但却造成了生产成本提高,以及土壤磷素异常累积、水体富营养化等问题(曹靖等,2000)。揭示磷高效植物基因型的形成机制,选育高磷效率基因型是开发利用土壤难溶态磷的有效技术途径。Nielsen等(1978)比较了玉米(Zea mays)近交种及其杂交种在田间对磷的吸收利用后指出玉米品种间磷积累量差异显著,且至少受2个基因控制。Mari等(2002;2003)研究发现在限制养分供给条件下西加云杉(Picea sitchensis)和挪威云杉(P. abies)苗木生长及N、P利用效率家系效应显著,受较强的遗传控制。由于磷素在土体中的移动性较差,主要借助于扩散方式迁移到根系表面,因此根系发育对植物磷效率有着决定性的影响,高磷效率基因型常通过改变根系形态来提高对土壤磷素的吸收和利用。如在缺磷条件下筛选出的磷高效蚕豆(Vicia faba)品系的最大特点就是根系发达(王惠珍等,2005),在低磷胁迫下马尾松(Pinus massoniana)高磷效率种源和家系皆表现出具有发达的须侧根(周志春等,2004b;2005)。研究养分胁迫环境下植物根系生长和形态参数的遗传特点和基因作用方式,对于营养高效基因型选育和新种质创制具有重要理论指导意义(王秀全等,2000)。
马尾松是我国松树中分布最广的树种,生长快,适应性强,与杉木(Cunninghamia lanceolata)并列为南方山地的两大针叶商品用材树种,广泛用于制浆造纸、各种板材、建筑、松香等,支撑着我国众多的造纸、木材加工、林产化工等产业发展。前期研究证实,马尾松吸收和利用土壤磷素的能力存在显著的种源和家系差异,其磷效率及相关子性状受中度以上的遗传控制,低磷胁迫下的侧根总长、侧根数、根体积等根系参数可很好地用于预测马尾松基因型的磷效率(周志春等,2004b; 2005)。本文在已有研究基础上,利用15个组合的马尾松测交系遗传交配设计双亲控制授粉子代种子,设置低磷和高磷2种磷素水平的盆栽试验,开展不同磷素水平下马尾松苗木生长和根系参数的配合力研究,揭示苗木生长和根系参数的遗传控制方式及与磷素环境的关系,为马尾松磷效率育种策略制定和高磷效率基因型选育提供科学理论依据。
1 材料与方法 1.1 试验材料试验材料来自1993—1994年在浙江省淳安县姥山林场马尾松一代育种群体中开展测交遗传交配设计获得的双亲控制授粉种子(金国庆等,2008)。基于优树子代测定结果,选择1116(广西)、1121(广西)和1126(广西)3个生长优良的无性系为父本,1134(广西)、1145(广西)、1201(贵州)、5915(浙江)和9236(福建)5个生长良好的无性系为母本开展测交设计的杂交制种,计有15个杂交组合。盆栽试验基质取自中国林业科学研究院亚热带林业研究所虎山的贫瘠酸性红壤,其有机质含量为6.11 g·kg-1,全N和全P含量分别为0.34 g·kg-1,0.33 g·kg-1,水解N、有效K和有效P含量分别为30.97 mg·kg-1,220.68 mg·kg-1和5.25 mg·kg-1,pH4.47。
1.2 试验设计基质土壤经风干过筛后与过磷酸钙、珍珠岩一起混合均匀后装盆,在中国林业科学研究院亚热带林业研究所温室大棚内开展盆栽试验。试验设置低磷(不施P)和高磷(土壤中过磷酸钙含量为1.5 g·kg-1)2种磷水平处理,20盆小区,共计600盆。试验用磷肥为浙江绍兴产,含P2O5 12.4%。营养杯直径8 cm、高10 cm,每营养杯装土约1.5 kg。2007年3月12日将种子直接点播营养杯中,每杯播种2粒,出苗后每杯保留1株,正常管理直到收获。整个试验过程中N、K等其他养分保持正常水平。
1.3 数据采集与统计分析试验于2007年10月中旬收获。不同磷肥处理下每杂交组合随机选取10株生长正常的苗木,量测苗高和地径后,先测定苗木主根长度,计数一级侧根数,并用加拿大REGENT公司生产RHIZO Pro STD1600+型根系图像分析系统测定根系总长、根总表面积和根系总体积等根系参数,然后将苗木分成根、茎、叶3部分,经105 ℃杀青30 min,80 ℃烘干至恒重,测定各部分的干物质量。按NCⅡ方法进行配合力分析,以单株数据进行配合力方差分析,按模型Ⅰ(固定模型)估算一般配合力(GCA)和特殊配合力(SCA)效应值等,按模型Ⅱ(随机模型)估算遗传方差分量和遗传力等(沈熙环,1990;朱军,1997;续九如,2006)。数据处理与统计分析在EXCEL和SAS软件中完成。
2 结果与分析 2.1 不同磷水平下苗木生长和根系参数的遗传差异和配合力方差分析表 1方差分析表明,不管在低磷还是在高磷水平下马尾松苗木生长和根系参数在组合间均达极显著差异,说明通过不同产地无性系间的杂交制种,可为下一世代育种创造变异丰富的基本群体,为创制和筛选优良杂交组合成为可能。较之于高磷水平,低磷水平下马尾松测交系苗木生长量和根系参数的平均值明显减小,但除干物质积累量外各性状的遗传变异系数都较大,如低磷水平苗高、侧根数、根体积的遗传变异系数分别为22.4%,18.8%和27.0%,分别为高磷水平的153.2%,126.2%和129.2%,说明在低磷胁迫下更易鉴别马尾松不同杂交组合间苗木生长和根系参数的遗传差异,筛选出耐低磷的优良杂交组合。
|
|
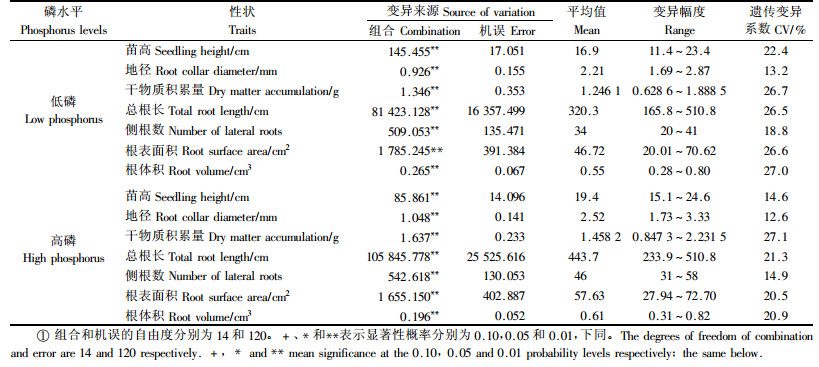
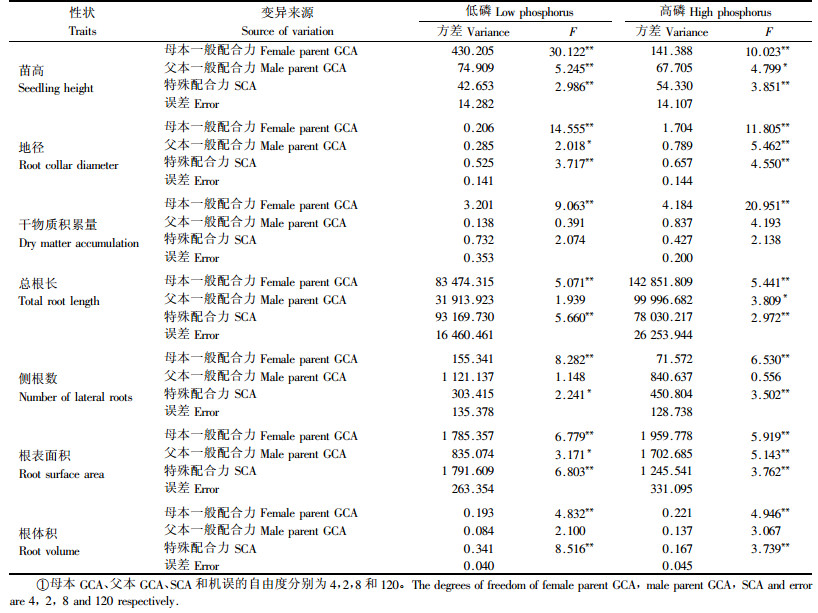
由于马尾松测交系苗木生长和根系参数差异显著,可进一步开展配合力方差分析。表 2的配合力方差分析结果表明,在低磷和高磷2种磷水平下,各生长和根系性状的母本GCA及SCA效应均达到显著和极显著水平。在2种磷水平下苗高、地径和根表面积的父本GCA效应显著或极显著,但除高磷下总根长外皆未发现干物质积累量、侧根数和根体积存在显著的父本GCA效应。
|
|
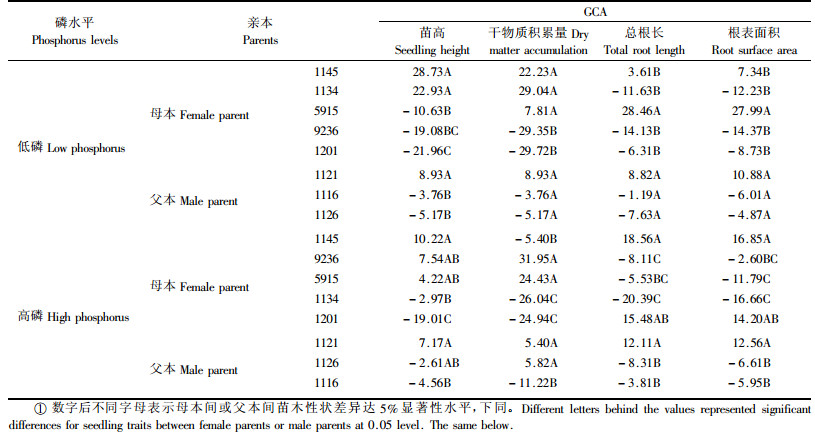
GCA是指某一亲本在许多杂交后代中的平均表现,可以反映其亲本的育种利用价值并能预测后代的表现(罗俊等,2004)。这里选择苗高、干物质积累量、总根长和根表面积4个重要性状开展GCA分析(表 3)。待测无性系(母本)GCA效应值的显著性测验表明,低磷水平下来自马尾松分布区南部广西1145和1134两个无性系的苗高和干物质积累量GCA最高,显著优于其他待测无性系,而来自贵州1201无性系表现最差;对于总根长和根表面积2个重要根系参数,以来自马尾松分布区北部浙江5915无性系的GCA最高,来自福建9236无性系最低,这一结论基本符合马尾松南部无性系苗木地上部分生长快、地下部分根量少,北部无性系根系发达、地上部分生长较慢的规律。随着磷素环境的改善,各待测无性系生长和根系参数的GCA效应值发生了明显的变化。1145和1201两个无性系苗高仍分别维持最大和最小的GCA效应值,但两者的总根长和根表面积GCA却从低磷时的中等值变成了高磷时的最高值。9236和5915两个无性系在低磷水平下苗高和干物质积累量GCA较小,施用磷肥后却显著提高了,两者干物质积累量的GCA最大。低磷水平下5915无性系的总根长和根表面积GCA最大,但在高磷水平下却显著降低。对于测交无性系(父本),不管在低磷还是在高磷下,生长和根系参数的GCA效应值都是1121无性系较大,1116无性系较小。这里还发现,待测无性系(母本)苗木生长和根系参数GCA间的差异远大于测交无性系(父本)GCA间的差异,这与待测无性系来自于不同产地有关。
|
|
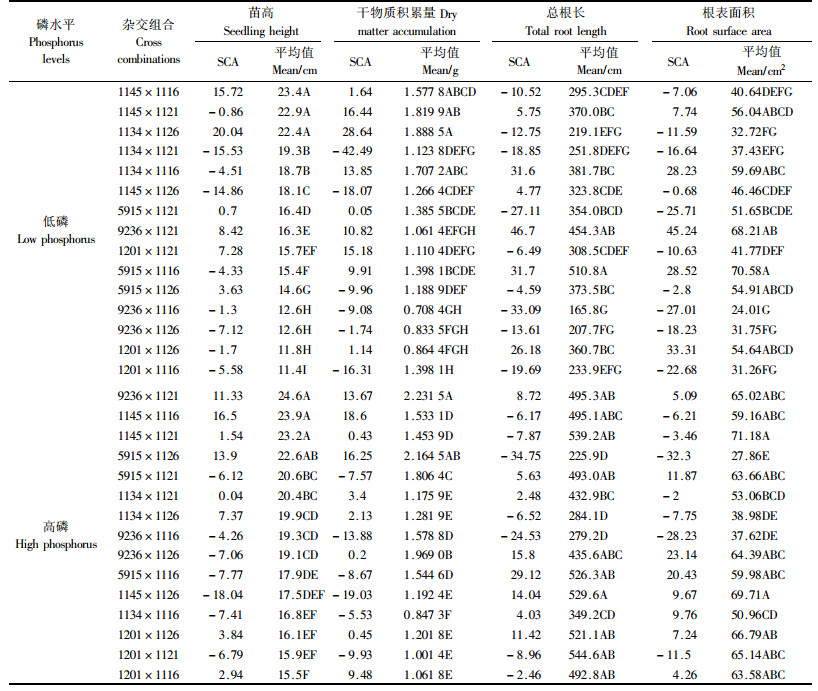
马尾松测交子代的性状表现既受亲本GCA效应影响,也受SCA效应制约。性状平均值是反映家系(组合)总体表现的参数,而SCA效应则是某一特定组合后代偏离双亲GCA的表型偏差。综合各杂交组合SCA效应值与平均值大小来看(表 4),在低磷水平下来自南部无性系间的杂交组合苗高生长量大,干物质积累量高,如1145×1116和1134×1126的苗高最大,1134×1121次之;1134×1126和1134×1116干物质积累量最高,1145×1121其次;对于总根长和根表面积2个根系参数,1134×1116、9236×1121和5915×1116较大。在高磷水平下,除上述1145×1116等个别南方无性系间的杂交组合生长仍表现较好外,其他苗高生长量大、干物质积累量高、根系发达的全同胞家系则来自中北部无性系与南部无性系间的杂交组合,如9236×1121和5915×1126等。可见各杂交组合生长和根系性状的SCA在不同磷素环境中表现有较大差异,说明磷素水平可以影响杂交组合特殊配合力的表现。
|
|
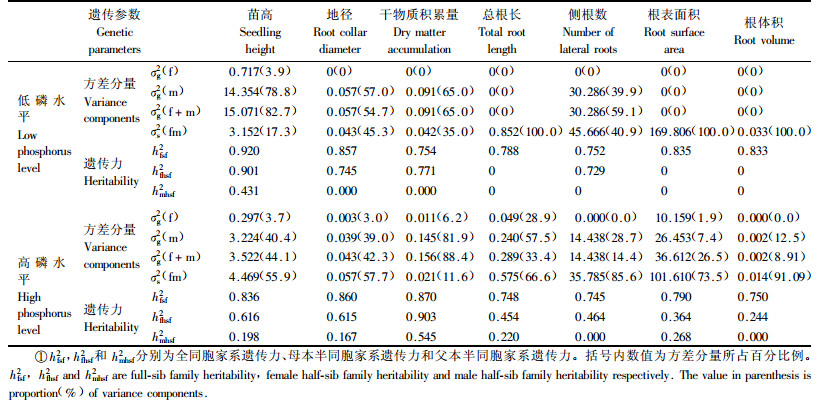
由表 5可知,不管是在低磷还是高磷水平下马尾松苗木生长和根系性状的母本GCA方差分量均显著地大于父本GCA方差分量,说明母本对测交子代生长的影响较大,父本效应相对较小。在高磷水平下,除单株干物质积累量外,苗高、地径、总根长、侧根数、根表面积和根体积的SCA效应均高于GCA效应,说明在磷素充足条件下马尾松苗木生长和根系性状主要受非加性遗传控制,加性基因效应则次之,尤其对于根系性状,GCA效应很小,仅为SCA效应的1/2至1/10,这与周志春等(2004a)和金国庆等(2008)已有研究结果是一致的,即杂交亲本若经过GCA测定,其子代性状的遗传控制以非加性基因效应起主导作用,加性基因效应次之。在低磷条件下,苗木总根长、根表面积和根体积3个根系参数仍表现出显著的SCA效应,GCA效应则可忽略。然而与高磷水平时比较,低磷水平时马尾松苗高、地径和侧根数的GCA效应显著提高,SCA效应明显减小,GCA效应对杂交组合苗高、地径和侧根数的贡献率分别为82.7%,54.65%和59.1%,说明在低磷水平下这3个性状的非加性基因效应变小了,较多地受加性遗传控制。这里还发现在2种磷处理条件下马尾松单株干物质积累量所受的遗传控制都以加性基因效应为主,不因磷素环境改变而异。表 5的性状遗传力估算结果显示,在低磷和高磷水平下马尾松生长和根系参数的全同胞家系遗传力都很高,其值在0.745~0.920,说明这些性状皆受强度的遗传控制,对全同胞家系选择可获得较高的遗传增益。此外不同磷水平下生长和根系参数的母本半同胞家系遗传力均大于父本半同胞遗传力,尤其苗高、地径、干物质积累量和侧根数的母本半同胞家系遗传力较高,其估算值变化在0.464~0.903。
|
|
研究结果表明,不管是在低磷还是在高磷水平下,马尾松测交系间苗木生长和根系性状皆存在显著的遗传差异。较之于父本GCA效应,母本GCA效应和SCA效应非常显著,测交系子代苗木生长和根系发育较多地受杂交母本及父母本互作的影响,通过不同产地无性系间的杂交可以选配强优势的杂种后代。本试验的待测无性系亲本是经过生长GCA初步测定的,在高磷或正常供磷条件下马尾松苗木生长和根系性状主要受非加性遗传控制,加性基因效应次之,尤其根系性状的加性基因效应很小。这一结果与齐明(1996)对杉木及周志春等(2004a)、金国庆等(2008)对马尾松幼林生长性状的研究结论一致,并进一步支持了Sprague等(1942)对玉米配合力育种的研究结论,即对于经过GCA测定的育种亲本,则需要进行SCA的选择与测定,创制亲本优势互补的杂交组合。马尾松南部无性系亲本地上部分生长量大、地下根系不发达,而北部无性系亲本地上部分生长量小、地下根系发达,其间的杂交组合如9236×1121和5915×1126等在高磷水平下表现出苗高生长量大、干物质积累量高、根系发达的特点。
低磷胁迫下马尾松测交系子代苗木生长和根系性状GCA/SCA相对重要性发生了明显的变化。苗高、地径和侧根数的GCA效应较高磷水平显著提高,SCA效应则明显减小。马尾松对低磷胁迫的应答涉及根系形态、根系分泌物、膜与体内磷转运以及菌根等许多生理过程,表现为数量遗传性状及受多基因控制。低磷胁迫时控制植物生长的数量遗传位点较磷供应充足时更多,推测这可能是低磷胁迫下马尾松生长性状GCA效应增加的内在原因之一。低磷水平下未发现马尾松苗木总根长、根表面积和根体积显著的父母本GCA效应,而父母本间的互作效应即SCA效应非常显著,这3个根系参数主要受显性等非加性基因效应控制。由于低磷胁迫下总根长、根体积等根系参数是预测马尾松种源和家系磷效率的特异性指标(周志春等,2004b;2005),可加强这些根系性状的特殊配合力育种,并在低磷条件下开展根系性状测试,以选育须侧根发达的高磷效率杂交组合。
曹靖, 张福锁. 2000. 低磷条件下不同基因型小麦幼苗对磷的吸收和利用效率及水分的影响. 植物生态学报, 24(6): 731-735. DOI:10.3321/j.issn:1005-264X.2000.06.015 |
金国庆, 秦国峰, 储德裕, 等. 2008. 马尾松测交系杂交子代生长性状遗传分析. 林业科学, 44(1): 70-76. DOI:10.3321/j.issn:1001-7488.2008.01.012 |
李庆逵. 1985. 中国红壤. 北京: 科学出版社, 145-146.
|
罗俊, 周会, 张木清, 等. 2004. 能源甘蔗主要经济和光合性状的遗传分析. 应用与环境生物学报, 10(3): 268-273. DOI:10.3321/j.issn:1006-687X.2004.03.002 |
齐明. 1996. 杉木育种中GCA与SCA的相对重要性. 林业科学研究, 9(5): 498-503. DOI:10.3321/j.issn:1001-1498.1996.05.010 |
沈熙环. 1990. 林木育种学. 北京: 中国林业出版社, 135-146.
|
王秀全, 陈光明, 何丹, 等. 2000. 玉米根系性状的遗传及相关分析. 中国农业科学, 33(增刊): 119-123. |
王惠珍, 张恩和, 彭忠. 2005. 不同春蚕豆品系对不同磷处理的适应机理研究. 草业学报, 14(2): 93-96. DOI:10.3321/j.issn:1004-5759.2005.02.016 |
续九如. 2006. 林木数量遗传学. 北京: 高等教育出版社.
|
周志春, 金国庆, 秦国峰, 等. 2004a. 马尾松纸浆材重要经济性状配合力及杂种优势分析. 林业科学, 40(4): 52-57. |
周志春, 谢钰容, 金国庆, 等. 2004a. 马尾松磷效率及其相关性状的家系遗传和变异. 北京林业大学学报, 26(6): 1-5. |
周志春, 谢钰容, 金国庆, 等. 2005. 马尾松种源磷效率研究. 林业科学, 41(4): 25-30. DOI:10.3321/j.issn:1001-7488.2005.04.005 |
朱军. 1997. 遗传模型分析方法. 北京: 中国农业出版社.
|
Mari S, Jonsson A, Thompson D, et al. 2002. Variation in nutrient utilization and juvenile growth in open-pollinated families of Picea sitchensis (Bong.) Carr. grown in a phytotron and correlations with field performance. SilvaeGenetica, 51(5/6): 225-232. |
Mari S, Jansson G, Jonsson A. 2003. Genetic variation in nutrient utilization and growth traits in Picea abies seedlings. Scandinavian Journal of Forest Research, 18(1): 19-28. DOI:10.1080/02827581.2003.10383134 |
Nielsen N E, Bareer S A. 1978. Differences among genotypes of corn in the kinetics of P uptake. Agronomy Journal, 70: 695-698. |
Sprague G F, Tatum L A. 1942. General vs specific combining ability in single crosses of corn. Journal American Society of Agronomy, 34(8): 923-932. |