 2009, Vol. 45
2009, Vol. 45文章信息
- 汤孟平, 周国模, 陈永刚, 赵明水, 何一波.
- Tang Mengping, Zhou Guomo, Chen Yonggang, Zhao Mingshui, He Yibo
- 基于Voronoi图的天目山常绿阔叶林混交度
- Mingling of Evergreen Broad-Leaved Forests in Tianmu Mountain Based on Voronoi Diagram
- 林业科学, 2009, 45(6): 1-5.
- Scientia Silvae Sinicae, 2009, 45(6): 1-5.
-
文章历史
- 收稿日期:2007-09-27
- 修回日期:2009-04-18
-
作者相关文章


2. 天目山国家级自然保护区管理局 临安 311300
2. Management Office, National Nature Reserve of Tianmu Mountain Lin'an 311300
传统方法常用混交比来反映混交林的树种空间隔离程度。但混交比仅能说明混交林各树种所占的比例,是一个非空间结构指数,并不能准确反映树种之间的空间隔离关系。Pielou(1961)曾提出用分离指数表示2个物种的个体分离情况(雷相东等,2002),但该指数对于多个种也只能进行两两比较(张金屯,1995)。鉴于此,Von Gadow等(1992)提出混交度的概念,即对象木的最近邻木中与对象木不属同种的个体所占的比例。混交度可以较好地表达树种空间隔离程度(惠刚盈等,2001)。
在混交度研究中,关于其最重要的参数即对象木的最近邻木株数n的取值尚存有争议。Füldner (1995)认为n=3。而惠刚盈等(2001)的研究指出,n=4可以满足对混交林空间结构分析的要求。实际上,n取值过大时,可能把非最近邻木也纳入计算范围;n取值过小时不能兼顾对象木周围最近邻木的所有可能情形。总之,这2种状况都将导致混交度的有偏估计。用Voronoi图确定最近邻木株数,则可以克服以上缺点。该方法由俄国数学家Voronoi在1908年提出,1911年荷兰气象学家Thiessen将其应用于气象观测(王新生等,2002)。Voronoi图是以诸多地理空间实体作为生长目标,按距离每一目标最近原则,将整个连续空间剖分为若干个Voronoi多边形,每一个Voronoi多边形只包含一个生长目标(陈军等,2003)。由于Voronoi图中的空间实体与Voronoi多边形一一对应,常用Voronoi多边形确定空间实体的影响范围(陈军,2002),如城市的吸引范围、竞争分析等(李新运等,2004;朱渭宁等,2004)。Voronoi图完全可以引入到混交度研究中,以确定对象木的最近邻木。因为,根据Voronoi图的特征可知,任意一株对象木的最近邻木是唯一确定的,由此计算的混交度准确可靠。
本研究以浙江天目山国家级自然保护区内的常绿阔叶林群落为例,对单株树木定位调查,用优势度分析方法确定群落优势种群,比较基于Voronoi图的混交度和传统混交度,分析种群间相互隔离的空间关系,为常绿阔叶林的植被保护、恢复与重建提供参考依据。
1 研究区概况浙江天目山国家级自然保护区位于浙江西北部临安市境内的西天目山(119°23′47″—119°28′27″E, 30°18′30″—30°24′55″N),总面积1 050 hm2。年均气温8.8~14.8 ℃;≥10 ℃年积温2 500~5 100 ℃;年降水量1 390~1 870 mm,相对湿度76%~81%。保护区地处中亚热带北缘向北亚热带过渡的地带,受海洋暖湿气候影响,温暖湿润,雨量充沛,森林植被十分茂盛。植被分布有明显的垂直界限,自山麓到山顶垂直带谱为:海拔870 m以下为常绿阔叶林;870~1 100 m为常绿、落叶阔叶混交林;1 100~1 380 m为落叶阔叶林;1 380~1 500 m为落叶矮林(汤孟平等,2007)。
2 研究方法 2.1 样地调查在保护区内,选择典型的常绿阔叶林设置1个大小为100 m×100 m的大样地,样地中心海拔630 m,主坡向南坡。用相邻格子调查方法,把样地划分为100个10 m×10 m的调查单元。在每个调查单元内,对胸径(DBH)≥5 cm的树木进行每木调查,采用激光对中全站仪(徕卡TCR702Xrange)测定三维坐标(X,Y,Z),并记录树种,测定每株树木的胸径、树高、枝下高和冠幅等因子。
2.2 优势种群的确定群落的优势种采用优势度分析法确定(Ohsawa,1984;达良俊等,2004),公式如下:
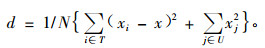
|
(1) |
式中:xi为前位树种(T)的相对基部面积(%);xj为剩余种(U)的百分比;N为总种数;x为按假设优势种个数确定的优势种理想百分比;d为各树种实际百分比与理想百分比之间的偏差,当偏差d达到最小值时,对应的前位树种即优势种。本研究中,相对基部面积就是胸高断面积百分比。前位树种是指按胸高断面积百分比由大到小排序,排列在最前面的为假设优势种。假设群落只有一个优势种,则优势种的理想百分比为100%;假设有2个优势种,则它们的理想百分比分别为50%;假设有3个优势种,则它们的理想百分比分别为33.3%;依次类推。
2.3 混交度计算混交度计算公式为(惠刚盈等,2001)
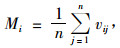
|
(2) |
式中:Mi为对象木i的混交度,n为最近邻木株数,vij是一个离散性的变量,当对象木i与第j株相邻木非同种时vij=1,反之,vij=0。
按(2)式计算的混交度是以对象木为中心的局部混交度。对林分还要计算平均混交度:
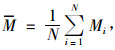
|
(3) |
式中:M为林分混交度,N为林分内林木株数,Mi为第i株树木的混交度。
关于(2)式中最近邻木株数n的取值,本研究采用惠刚盈等(2001)对传统混交度的研究结果n=4和基于Voronoi图的相邻多边形个数2种方法进行比较分析。
Voronoi图由一组连续多边形组成(邬伦等,2001)。每株树木被视为平面上的一个点,由树木点位置生成Voronoi图。根据Voronoi图的特征,每个Voronoi多边形内仅包含1株树木。显然,对象木的最近邻木株数与相邻Voronoi多边形的个数相等(图 1)。为便于区别,把惠刚盈等(2001)取n=4的传统混交度记为M,把基于Voronoi图的混交度记为MV。

|
图 1 基于Voronoi图的最近邻木 Figure 1 Nearest tree based on Voronoi diagram |
当对象木到最近邻木的距离大于它到样地边界的最近距离时,必须进行边缘校正(吴巩胜等,2000):本研究采用平移式八邻域大样地边缘校正法。当采用Voronoi图确定对象木的最近邻木时,还需要生成以大样地内每株树木为生长目标的Voronoi图。应当说明,本研究只把原样地中胸径≥5 cm的树木作为对象木,并计算其混交度。
3 结果与分析 3.1 MV与M的关系利用调查的1 603株树木的混交度MV和M绘制散点图,并拟合关系曲线(图 2)。由图 2可见,MV和M有较高的相关性,说明采用MV和M描述物种相互隔离程度具有一致性,均为有效混交度指数。但M只有5种可能取值0,0.25,0.5,0.75和1,难以描述复杂多样的种间隔离关系,这是n=4的限定性条件决定的。而MV并不固定n的取值,n是基于Voronoi图根据每株对象木周围最近邻木的实际情况确定的。本研究中,基于Voronoi图确定n的取值为3~13,共有11种可能取值,多数取值为5,6和7,平均取值为6(图 3)。

|
图 2 M与MV的关系 Figure 2 Relation between M and MV |

|
图 3 基于Voronoi图的对象木的最近邻木株数分布 Figure 3 Nearest tree number distribution of object tree based on Voronoi diagram |
图 4表现出M和MV与胸径的关系有共同趋势。当胸径<20 cm时,混交度M和MV取值在0~1之间;当胸径>20 cm后,混交度有增大趋势,且取值范围变窄。当胸径>50 cm后,混交度都等于1。总体来看,随胸径增大,混交度增大。Moeur等(1993)研究证实,在群落中大树呈均匀分布,小树呈聚集分布。由于均匀分布的大树降低了同种接触的可能性,从而提高了大树的混交度。小树的混交度则取决于种群繁殖方式和种间关系,因而表现为混交度的取值范围较宽。

|
图 4 混交度与胸径的关系 Figure 4 Relation between mingling and DBH |
常绿阔叶林的物种丰富,结构复杂。采用(1)式确定群落优势种群,并进行混交度比较分析。根据调查统计,样地内胸径≥5 cm的树木有1 603株,共73个树种,其中优势种群11个(表 1)。
|
|

各优势种群的样地平均混交度见图 5。MV和M十分接近,但除杉木(Cunninghamia lanceolata)和枫香(Liquidambar formosana)外,M一般都小于MV, 说明M低估了树种相互隔离程度。

|
图 5 优势种群混交度 Figure 5 Mingling of dominant populations |
样地平均混交度M=0.720 7,MV=0.743 1(图 5),表明天目山自然保护区常绿阔叶林的混交度较大,不同树种之间的相互隔离程度较高。除细叶青冈(Cyclobalanopsis myrsinaefolia)、青冈(C.glauca)和短尾柯(Lithocarpus brevicaudatus)外,其他各优势种群的混交度都大于样地平均混交度,说明这些优势种群不仅胸高断面积占优势,而且树种之间相互隔离程度也较高。而细叶青冈、青冈和短尾柯的混交度小于样地平均混交度,主要是因为这3个树种属壳斗科(Fagaceae)植物,具有实生和萌生2种聚集繁殖方式(胡小兵等,2003),从而降低了物种隔离程度。
3.4 优势种群与非优势种群混交度比较图 6表明,优势种群比非优势种群的平均混交度略低。原因是在优势种群中,丛生性较强、混交度偏低的细叶青冈、青冈和短尾柯的株数占优势种群的67.65%,占群落的52.71%。无论是优势种群还是非优势种群,与MV相比,M低估了树种的相互隔离程度。

|
图 6 优势种群与非优势种群混交度 Figure 6 Mingling of dominant populations and non-dominant populations |
常绿种群是常绿阔叶林的标志性成分,对群落物种结构有重要影响。图 7显示,常绿种群与非常绿种群的混交度都比较高。但常绿种群比非常绿种群的平均混交度略低。这同样是由于在整个群落中占优势的常绿树种细叶青冈、青冈和短尾柯等具有聚集分布现象(汤孟平等,2006),也可以看出,无论是常绿种群还是非常绿种群,M都小于MV。

|
图 7 常绿种群与非常绿种群的混交度 Figure 7 Mingling of evergreen population and non-evergreen population |
基于Voronoi图的混交度MV和传统混交度M有较高的相关性和一致的趋势,均为有效混交度指数。但MV通过Voronoi图的相邻多边形确定对象木的实际最近邻木,比混交度M更准确地描述了种间相互隔离关系。尽管MV和M十分接近,但一般地,M小于MV,M往往低估了实际混交度,且M取值缺少中间状态,导致M的变动幅度大于MV。可见,MV是优于M的混交度指数。
常绿阔叶林群落中,单株树木的混交度随胸径的增加而增大,当胸径<20 cm时,混交度的取值为0~1,范围最宽,当胸径>20 cm后,混交度有增大趋势,当胸径>50 cm后,混交度都等于1。
常绿阔叶林群落的平均混交度M=0.720 7,MV=0.743 1,表明该群落不同树种之间的相互隔离程度较高。群落的11个优势种群中,多数优势种群的混交度都大于群落平均混交度,少数优势种群包括细叶青冈、青冈和短尾柯的混交度低于群落平均混交度。具有聚集繁殖方式的壳斗科植物细叶青冈、青冈和短尾柯既是优势种群,又是常绿种群,导致优势种群的平均混交度低于非优势种群和常绿种群的平均混交度低于非常绿种群的平均混交度。
壳斗科植物的聚集繁殖方式不仅加剧了青冈等常绿优势种群的种内竞争,而且还降低了其种群相互隔离程度。也正是这种繁殖方式使青冈等常绿树种在常绿阔叶林中确立了牢固的优势地位,对群落演替产生了重要影响。因此,在常绿阔叶林经营或植被恢复与重建过程中,既不能忽视种群的生物学特性,盲目追求混交度等于1的最高混交状态,也不能忽视种间关系,任种群自由繁殖和发展。以常绿优势种群为关键切入点,进一步研究常绿阔叶林种群繁殖与调控机制是构建常绿阔叶林群落理想空间结构的基本前提。
基于Voronoi图的混交度研究需要利用GIS的相关空间分析功能。现有GIS软件如MapInfo、ArcMap等都有现成的Voronoi图分析功能。因此,这种空间分析功能很容易实现。实际上,此空间分析功能也可应用于研究其他群落尺度的林学或生态学问题如物种竞争、林木空间分布格局等。
陈军, 赵仁亮, 乔朝飞. 2003. 基于Voronoi图的GIS空间分析研究. 武汉大学学报:信息科学版, 28(增): 32-37. |
陈军. 2002. Voronoi动态空间数据模型. 北京: 测绘出版社.
|
达良俊, 杨永川, 宋永昌. 2004. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型. 植物生态学报, 28(3): 376-384. DOI:10.3321/j.issn:1005-264X.2004.03.013 |
胡小兵, 于明坚. 2003. 青冈常绿阔叶林中青冈种群结构与分布格局. 浙江大学学报:理学版, 30(5): 574-579. |
惠刚盈, 胡艳波. 2001. 混交林树种空间隔离程度表达方式的研究. 林业科学研究, 14(1): 23-27. DOI:10.3321/j.issn:1001-1498.2001.01.004 |
雷相东, 唐守正. 2002. 林分结构多样性指标研究综述. 林业科学, 38(3): 140-146. DOI:10.3321/j.issn:1001-7488.2002.03.025 |
李新运, 郑新奇. 2004. 基于曲边Voronoi图的城市吸引范围挖掘方法. 测绘学院学报, 21(1): 38-41. |
汤孟平, 周国模, 施拥军, 等. 2006. 天目山常绿阔叶林优势种群及其空间分布格局. 植物生态学报, 30(5): 743-752. DOI:10.3321/j.issn:1005-264X.2006.05.004 |
汤孟平, 周国模, 施拥军, 等. 2007. 不同地形条件下群落物种多样性与胸高断面积的差异分析. 林业科学, 43(6): 27-31. |
王新生, 李全, 郭庆胜, 等. 2002. Voronoi图的扩展、生成及其应用于界定城市空间影响范围. 华中师范大学学报:自然科学版, 36(1): 107-111. |
吴巩胜, 王政权. 2000. 水曲柳落叶松人工混交林中树木个体生长的竞争效应模型. 应用生态学报, 11(5): 646-650. DOI:10.3321/j.issn:1001-9332.2000.05.002 |
邬伦, 刘瑜, 张晶, 等. 2001. 地理信息系统——原理、方法和应用. 北京: 科学出版社, 201-202.
|
张金屯. 1995. 植被数量生态学方法. 北京: 中国科学技术出版社, 87-89.
|
朱渭宁, 马劲松, 黄杏元, 等. 2004. 基于投影加权Voronoi图的GIS空间竞争分析模型研究. 测绘学报, 33(2): 146-150. DOI:10.3321/j.issn:1001-1595.2004.02.010 |
Füldner K. 1995. Strukturbeschreibung von buchen-edellaubholz-mischwldern. Doctoral dissertation, University of Gttingen.
|
Moeur M. 1993. Characterizing spatial patterns of trees using stem-mapped data. Forest Science, 39(4): 756-775. |
Pielou E C. 1961. Segregation and synmmetry in two species populations as studied by nearest neighbour relations. Ecology, 49: 255-269. DOI:10.2307/2257260 |
Ohsawa M. 1984. Differentiation of vegetation zones and species strategies in the subalpine region of Mt. Fuji. Vegetation, 57: 15-52. DOI:10.1007/BF00031929 |
von Gadow K, Füldner K. 1992. Zur Methodik der Bestandesbeschreibung. Klieken: Vortrag anlässlich der Jahrestagung der AG Forsteinrichtung.
|