 2009, Vol. 45
2009, Vol. 45文章信息
- 李永红, 马颖敏, 韩蕾.
- Li Yonghong, Ma Yingmin, Han Lei
- 超氧化物歧化酶与马拉巴栗种子脱水耐性之间的关联
- Relationship between the Tolerance to Dehydration and Superoxide Dismutases (SODs) in Pachira macrocarpa
- 林业科学, 2009, 45(5): 74-79.
- Scientia Silvae Sinicae, 2009, 45(5): 74-79.
-
文章历史
- 收稿日期:2008-05-28
-
作者相关文章



2. 中国林业科学研究院林业研究所 国家林业局林木培育重点实验室 北京 100091
2. Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration Research Institute of Forestry, CAF Beijing 100091
马拉巴栗(Pachira macrocarpa),也叫台湾发财树,木棉科(Bombacaceae)瓜栗属。生存于热带雨林地区,原产墨西哥和哥斯达黎加等地,是著名的观叶植物。马拉巴栗种子多胚,1粒种子可发1~3芽,通常播种育苗来繁殖新株。据调查,马拉巴栗种子成熟后不耐保存,1周左右种子活力便急速下降,且该种子也不耐脱水,当脱水至20%时,50%以上的种子会丧失活力,目前未有贮藏马拉巴栗种子的理想方法。本研究以马拉巴栗种子为对象,研究种子脱水耐性的机制,旨在为探索适宜马拉巴栗种子的贮藏方法提供理论依据。
对豌豆(Pisum sativum)(Alscher et al., 2002; Gomez et al., 2004)、白桦(Betula platyphylla)(孙国荣等,2003)、百脉根(Lotus corniculatus)(Borsani et al., 2001)、月季(Rosa hybrida)(单宁伟等,2005)、百合(Lilium)(尹慧等, 2007)、叶底珠(Securinega suffruticosa)(徐兴友等, 2008)的研究发现,失水胁迫会诱导细胞中SOD活性增高,导致SOD同工酶条带发生变化。但迄今为止,有关马拉巴栗种子脱水耐性与SOD同工酶的相关性研究未见报道。因此,本研究拟从SOD活性和同工酶水平上明确马拉巴栗种子失水胁迫耐性与其SOD之间的关联。
1 材料与方法 1.1 材料及处理2007年8月在深圳市四季青花场,采收因成熟而自然脱落的果实内的种子。种子采收后0.5 h内运回深圳职业技术学院应用化学与生物技术学院实验室,进行试验。
1.2 方法将种子分别浸泡于0,5,10,20,30,40,50 mmol·L-1二乙基二硫代氨基甲酸(diethyldithiocarbamate, DDTC,SOD专一性抑制剂)(Bailly et al., 2001)溶液中,以室温下放置的种子作对照。在处理中,每10粒种子用100 mL液体浸泡。在室温下以60 r·min-1的速度振摇吸收12 h。取出后在温度24~26 ℃下置于硅胶干燥器中10 h作脱水处理,种子与硅胶比例为1:5。然后取出测定种子含水量、相对电导率、MDA含量、SOD活性和种子活力指数。并通过这些结果,从中筛选出有效的DDTC预处理浓度;而后以此浓度和清水预处理种子12 h,再分别进行0,3,6,12,20 h的硅胶脱水,测定种子含水量、相对电导率、MDA含量和种子活力指数,并进行SOD同工酶凝胶电泳分析。每试验重复进行3次。
种子出苗率测定方法为每个处理播种50粒种子,10天后统计所有种子成苗的数量;种子活力指数测定参考吕成群等(2000)的方法;种子含水量测定参照杨期和等(2007)的方法;相对电导率和MDA含量参考林植芳等(1984)的方法,稍加改动。电导率用DDS-11A型电导率仪测定,用蒸馏水和去离子水冲洗干净后浸泡于100 mL去离子水中,加盖浸泡过夜后测定电导率R1,然后转至沸水浴中煮1 h,冷却至室温后测定电导率R2,按照相对电导率(%)=R1/R2×100%进行计算;MDA含量的测定,用每克材料加入10 mL 5%TCA的比例提取种子MDA,提取液离心后加入0.67%TBA 2 mL,100 ℃水浴中煮沸30 min后,分别测定450,532,600波长下的吸光值,按公式算出MDA含量。
SOD酶活性测定参照Van Breusegem等(1999)的方法稍加改动,以可抑制NBT光还原反应50%的酶量为1个SOD活性单位(1 unit);同工酶电泳采用不连续的Tris-甘氨酸聚丙烯酰胺凝胶垂直板装置电泳,染色采用改良苯胺法(廖兆周,1988)。
2 结果与分析 2.1 DDTC预处理液适宜浓度筛选 2.1.1 对种子萌发率和活力指数的影响由图 1可知,经过10 h硅胶干燥处理的种子萌发率和活力指数明显下降,且种子活力指数下降幅度更明显。不同处理间种子活力指数的差异显著(图 1B),种子萌发率和活力指数先迅速下降,后上升,以20 mmol·L-1DDTC处理的种子萌发率和活力指数最低,萌发率只有36%,比CK下降了64%;活力指数下降至52.06,比CK下降近90%。

|
图 1 不同浓度DDTC处理对种子干燥10 h后萌发率(A)和活力指数(B)的影响(n=3) Figure 1 Effect of different concentration of DDTC on seed germination rate (A) and seed vigor index (B) after 10 h dehydration (n=3) 均值±标准误Mean±SE。不同字母表示不同脱水处理之间差异显著(P < 0.05)。The different letters indicate significant difference at P < 0.05.下同。The same below. |
从图 2可知,经干燥处理后,种子相对电导率和MDA含量随DDTC浓度升高呈现先增加后降低的趋势,且以20 mmol·L-1处理为最大值。

|
图 2 不同浓度DDTC处理对种子干燥10 h后相对电导率和MDA含量的影响(n=3) Figure 2 Effect of different concentration of DDTC on seed electrical conductivity and MDA content after 10 h dehydration(n=3) |
从图 3可知,刚采收的新鲜种子SOD活力是281.087 U·g-1,经DDTC预处理后,除5,10 mmol·L-1 DDTC处理过的种子的SOD活性有所提高外,其他浓度处理后SOD活性均不同程度下降。与0 mmol·L-1DDTC处理的种子相比,其他浓度DDTC处理均明显使种子SOD活性受到不同程度的抑制,20 mmol·L-1 DDTC预处理种子中的SOD活性最低,下降幅度为78.83%。

|
图 3 不同浓度DDTC处理对种子干燥10 h后SOD活性的影响(n=3) Figure 3 Effect of different concentration of DDTC on SOD specific activity after 10 h dehydration(n=3) |
综上所述,20 mmol·L-1DDTC预处理比较成功,有效地抑制了成熟马拉巴栗种子的SOD活性,使硅胶干燥后的相对电导率和MDA含量明显增高,种子活力显著下降。因此,选用20 mmol·L-1DDTC对马拉巴栗种子进行不同干燥程度的研究。
2.2 DDTC预处理对种子脱水后活力的影响 2.2.1 硅胶脱水不同时间下种子含水量的变化如图 4所示,随着脱水的进行,种子含水量逐步降低。无论是否经过DDTC处理,材料在脱水初期的含水量下降较快,在脱水进行6 h以后,种子含水量的下降速度逐渐减缓。随着脱水的进行,种皮逐渐失去柔软性,变得脆硬,在脱水15 h左右,胚因为较大程度失水萎缩,与种皮明显分离。

|
图 4 20 mmol·L-1DDTC预处理的种子经不同程度硅胶脱水后的含水量变化(n=10) Figure 4 Changes of seed water content after 20 mmol·L-1DDTC pretreatment and dehydration(n=10) |
从图 5可以看出,种子的活力指数随着脱水时间的延长而降低。未经DDTC处理的种子脱水3 h后,种子活力指数下降到对照的85.05%,出苗率接近对照水平;经过DDTC处理3 h后,种子活力指数下降明显,为对照的47.09%,出苗率从202%下降到140%;未经DDTC处理的种子脱水20 h,种子活力指数迅速下降到对照的12.54%,出苗率也下降到54%;但是经过DDTC处理的种子,在脱水12 h后,种子活力指数下降到对照的11.98%,脱水20 h,种子活力仅为对照的3.90%,出苗率仅有26%。由此可见,马拉巴栗种子的活力与脱水程度有密切的关系,20 mmol·L-1DDTC预处理更进一步加剧了脱水对种子的活力指数和出苗情况的影响,整体种子活力急速下降。

|
图 5 20 mmol·L-1DDTC预处理的种子经不同程度硅胶脱水后种子萌发率、活力指数和出苗率的变化(n=3) Figure 5 Changes of seed germination rate, seed vigor index and germination rate after 20 mmol·L-1DDTC pretreatment and dehydration (n=3) |
从图 6可知,2种处理方式在3~20 h的脱水过程中,种子相对电导率和MDA含量呈现相同的变化趋势,即随脱水程度加重,两者均呈增加趋势,但是,相同的脱水时间,与对照(蒸馏水浸泡后直接脱水)相比,20 mmol·L-1DDTC预处理进一步加剧了种子相对电导率和MDA含量的增加,而且在脱水6 h后,经DDTC处理的与对照的差异更为明显。脱水6 h时,DDTC预处理的种子相对电导率是对照的298.5%,MDA含量是对照的225.2%,到脱水20 h(此时种子含水量为16.48%)时,DDTC预处理的种子相对电导率是对照的778%,MDA含量是对照的505.1%。

|
图 6 20 mmol·L-1DDTC预处理的种子经不同程度硅胶脱水后相对电导率和MDA含量的变化(n=3) Figure 6 Changes of seed relative electrical conductivity and MDA content after 20 mmol·L-1DDTC pretreatment and dehydration (n=3) |
如图 7所示,随着脱水的进行,SOD活性先轻微下降,随后上升,到脱水6 h以后,活性大幅下降。经过20 mmol·L-1DDTC预处理的种子在脱水20 h后,活性更降低至25.31 U·g-1。

|
图 7 20 mmol·L-1DDTC预处理的种子经不同程度硅胶脱水后SOD活性的变化(n=3) Figure 7 Changes of SOD specific activities after 20 mmol·L-1 DDTC pretreatment and dehydration(n=3) |
基于SOD不同类型同工酶对SDS、H2O2和KCN的不同反应,即MnSOD对KCN和H2O2都不敏感,但对SDS敏感;FeSOD对H2O2敏感,对1%SDS和KCN不敏感;Cu/ZnSOD对KCN和H2O2都敏感,但对1%SDS不敏感等特征,对马拉巴栗新鲜的成熟种子中SOD同工酶的类型进行了鉴定(图 8)。

|
图 8 SOD同工酶的鉴定 Figure 8 Identification of SOD isoenzymes A.未使用抑制剂染色Without inhibitors;B.活性染色前在预处理液中添加了3 mmol·L-1的KCN Preincubating the gels with 3 mmol·L-1 KCN;C.活性染色前在预处理液中添加了5 mmol·L-1H2O2 Preincubating the gels with 5 mmol·L-1H2O2;D.活性染色前在预处理液中添加了1%SDS的胶片Preincubating the gels with 1%SDS. |
图中B、C、D中的白框分别是凝胶预稳浴液加入KCN、H2O2和SDS后被抑制的SOD同工酶种类,根据H2O2、KCN和1%SDS对3种同工酶的不同抑制效应,鉴定出种子中MnSOD、FeSOD和Cu/ZnSOD这3种类型都存在。其中MnSOD有2种,FeSOD有4种,Cu/ZnSOD有2种;迁移率大小依次是Cu/ZnSOD>FeSOD>MnSOD;谱带强弱依次为FeSOD>Cu/ZnSOD>MnSOD。
分析种子中各类SOD同工酶相应种类的变化,可发现SOD只是谱带强弱上存在变化。
2.3.2 DDTC预处理对种子SOD同工酶的影响从图 9可知,与没有经过硅胶脱水的种子相比,经不同时间脱水处理后,种子SOD同工酶条带的亮度大部分都出现了变化,具体表现为:脱水3 h,除了Cu/ZnSOD2的亮度稍降外,8条条带亮度有不同程度的增强,MnSOD2及FeSOD2,FeSOD3,FeSOD4相对明显;脱水6 h,所有条带的亮度开始下降,FeSOD1,FeSOD4和Cu/ZnSOD2变得非常微弱;继续脱水至20 h,除了FeSOD3和Cu/ZnSOD1还留有较清晰条带外,其他条带基本消失。

|
图 9 脱水处理对种子SOD同工酶的影响 Figure 9 SOD isoforms with dehydration treatment |
图 10表明,经过20 mmol·L-1DDTC低温预处理的样品,与未经预处理的(图 9)相比,种子中各同工酶条带亮度明显降低,特别是FeSOD3的亮度下降最明显;经过脱水处理3~12 h后,除了MnSOD1,FeSOD3,Cu/ZnSOD1还能较清晰看到,其他条带全部消失。脱水至20 h,所有条带都消失。

|
图 10 DDTC处理后的脱水对种子SOD同工酶的影响 Figure 10 SOD isoforms after 20 mmol·L-1DDTC pretreatment and dehydration treatment |
由此推测,MnSOD,FeSOD1,FeSOD2,FeSOD4和Cu/ZnSOD2是与种子脱水耐性密切相关的条带。
3 讨论本研究在DDTC浓度筛选基础上,选择20 mmol·L-1的浓度有效地抑制了马拉巴栗种子SOD活性,使SOD活性从314.08 U·g-1急剧下降至66.501 U·g-1,抑制率达到78.83%。同时,20 mmol·L-1DDTC预处理加重了种子脱水过程中相对电导率、MDA含量的增加,明显降低了种子活力指数、出苗率,显著缩短了种子干燥保存过程中的寿命。上述研究结果证明种子中SOD活性降低后,种子的脱水耐性明显减弱,同时表明马拉巴栗种子脱水耐性伴随有SOD活性变化。通过相关分析显示,马拉巴栗种子含水量和种子萌发率、出苗率呈极显著正相关(r=0.997**; r=0.987**); 含水量与种子活力指数也呈显著正相关(r=0.941*);马拉巴栗种子萌发率与相对电导率和MDA含量彼此间呈负相关,相对电导率与萌发率之间达到极显著水平(r=-0.977**),MDA含量和萌发率之间也达到显著水平(r=-0.955*);种子出苗率与MDA含量和相对电导率之间也呈显著负相关(r=-0.916*;r=-0.948*);而活力指数与相对电导率、MDA含量之间虽然未达到显著水平,但相关系数均较高(r=-0.851;r=-0.829)。
从已有报道(Gomez et al., 2004; Borsani et al., 2001; 单宁伟等, 2005)分析,失水胁迫对SOD同工酶的影响因植物种类、胁迫强度等不同而异。例如,营养生长末期的豌豆(Pisum sativum cv. Linclin)苗经轻度盐胁迫后,Cu/ZnSOD增强,MnSOD活性没有变化;重度胁迫后Cu/ZnSODⅡ明显减弱,而FeSOD条带明显增强(Inaki et al., 1998)。百脉根水培苗经2~6 h失水胁迫后,叶片Cu/ZnSOD活性上升,MnSOD活性下降,FeSOD活性没有变化(Borsani et al., 2001)。切花月季Samantha花朵经失水胁迫处理后,诱导产生了3条新的FeSOD(FeSOD4~6)条带,同时MnSOD消失(单宁伟等, 2005)。
本研究表明,马拉巴栗成熟种子中MnSOD、FeSOD和Cu/ZnSOD都存在,其中,MnSOD有2种,FeSOD有4种,Cu/ZnSOD有2种。硅胶脱水处理后,只留下FeSOD和Cu/ZnSOD的部分条带(FeSOD3和Cu/ZnSOD1),其他条带消失,没有诱导产生新的条带。DDTC预处理再进行脱水处理,最后所有条带全部消失。基于上述结果可以初步推测马拉巴栗种子脱水过程中,有SOD同工酶带与活性变化与其相伴随。初步脱水,SOD同工酶条带亮度都明显增强,随着脱水程度加深,SOD同工酶条带亮度降低甚至部分消失,说明脱水过程中,SOD首先被诱导合成,致使其活性增强,脱水至一定程度后,SOD活性下降,这与SOD活性测定结果相吻合。
而经过DDTC预处理,种子中的SOD活性被抑制,加速了种子脱水过程的衰老和死亡。推测,SOD被抑制后,种子内产生的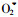
廖兆周. 1988. 甘蔗、斑茅及其杂种的过氧化物酶分析. 植物学报, 30(2): 214-219. |
林植芳, 李双顺, 林桂珠. 1984. 水稻叶片衰老与超氧化物歧化酶活性的关系. 植物学报, 26(6): 605-615. |
吕成群, 黄宝灵, 牟继平. 2000. 稀土对八角种子萌发及幼苗生长的生理效应. 广西科学, 7(3): 228-231. DOI:10.3969/j.issn.1005-9164.2000.03.019 |
单宁伟, 金基石, 赵喜亭, 等. 2005. 切花月季Samantha失水胁迫耐性与其超氧化物歧化酶之间的关联. 中国农业科学, 38(7): 1431-1438. DOI:10.3321/j.issn:0578-1752.2005.07.024 |
孙国荣, 彭永臻, 阎秀峰, 等. 2003. 干旱胁迫对白桦实生苗保护酶活性及脂质过氧化的影响. 林业科学, 39(1): 165-167. DOI:10.3321/j.issn:1001-7488.2003.01.027 |
徐兴友, 王子华, 张风娟, 等. 2008. 干旱胁迫对6种野生耐旱花卉幼苗根系保护酶活性及脂质过氧化作用的影响. 林业科学, 44(2): 41-47. DOI:10.3321/j.issn:1001-7488.2008.02.007 |
杨期和, 袁培华, 廖富林, 等. 2007. 假槟榔种子催芽技术和脱水耐性的研究. 广西植物, 27(6): 913-917. DOI:10.3969/j.issn.1000-3142.2007.06.021 |
尹慧, 陈秋明, 何秀丽, 等. 2007. 短暂高温对百合植株抗氧化酶系统的影响. 园艺学报, 34(2): 509-512. DOI:10.3321/j.issn:0513-353X.2007.02.049 |
Alscher R G, Erturk N, Heath L S. 2002. Role of superoxide dismutase (SODs) in controlling oxidative stress in plants. Journal of Experimental Botany, 53(372): 1331-1341. DOI:10.1093/jexbot/53.372.1331 |
Bailly C, Corbineau F, van Doom W G. 2001. Free radical scavenging and senescence in Iris tepals. Plant Physiology and Biochemistry, 39: 649-656. DOI:10.1016/S0981-9428(01)01289-X |
Borsani O, Diaz P, Agius M F, et al. 2001. Water stress generates an oxidative stress through the induction of a specific Cu/Zn superoxide dismutase in Lotus corniculatus leaves. Plant Science, 161: 757-763. DOI:10.1016/S0168-9452(01)00467-8 |
Gomez J M, Jimnez A, Olmos E, et al. 2004. Location and effects of long-term NaCl stress on superoxide dismutase and ascorbate peroxidase isoenzyme of pea (Pisum sativum cv. Puget)chloroplasts. Journal of Experimental Botany, 55(394): 119-130. |
Inaki I O, Pedro R E, Cesar A I, et al. 1998. Oxidative damage in pea plants exposed to water deficit or parquet. Plant Physiology, 116: 173-181. DOI:10.1104/pp.116.1.173 |
Van Breusegem F, Slooten L, Stassart J, et al. 1999. Effects of overproduction of tobacco MnSOD in maize chloroplast on foliar tolerance to cold and oxidative stress. Journal of Experimental Botany, 50(330): 71-78. DOI:10.1093/jxb/50.330.71 |