 2009, Vol. 45
2009, Vol. 45文章信息
- 臧润国, 刘华, 张新平, 丁易, 江晓珩, 郭仲军, 张炜银.
- Zang Runguo, Liu Hua, Zhang Xinping, Ding Yi, Jiang Xiaoheng, Guo Zhongjun, Zhang Weiyin
- 天山中部天然林分中不同龄级天山云杉光合特性
- Photosynthetic Characteristics of Picea schrenkiana var. tianschanica of Different Age Classes in a Natural Stand in the Central Part of Tianshan Mountains
- 林业科学, 2009, 45(5): 60-68.
- Scientia Silvae Sinicae, 2009, 45(5): 60-68.
-
文章历史
- 收稿日期:2008-03-01
-
作者相关文章


2. 安徽农业大学林学与园林学院 合肥 230036;
3. 新疆林业科学研究院森林生态研究所 乌鲁木齐 830000
2. School of Forestry & Landscape Architecture, Anhui Agricultural University Hefei 230036;
3. Institute of Forest Ecology, Xinjiang Academy of Forestry Urumqi 830000
近几十年来,有关光合作用的研究已经在从基因到生物圈的不同层次上开展,其中叶片水平上的光合作用体现了植物本身的生物学特性,它不仅为植物其他的生理活动提供能量和结构性物质,还可为个体和群体光合——生产力模型,乃至区域和全球模型的构建提供基础(Wilson et al., 2001;Lusk et al., 2003;Nedbal et al., 2007)。大部分针叶林分布在高纬度和高海拔的较贫瘠或干旱的地区(Becker, 2000)。云杉(Picea)为耐阴性较强的树种,在高纬度的寒带、寒温带至低纬度的暖温带与亚热带的亚高山与高山的阴坡、半阴坡和谷地形成纯林,或与冷杉(Abies)、落叶松(Larix)、铁杉(Tsuga)及阔叶树等组成针叶混交林或针阔混交林。目前,对云杉的光合生理生态特性研究包括云杉针叶的结构及其光捕获能力(Portsmuth et al., 2007)和光环境变化对云杉光合特性的影响等方面。如冠层结构对针叶生长所需光环境的影响(Stenberg et al., 1999;de Chantal et al., 2003;Niinemets, 2007)、野外遮荫条件下云杉阴生叶和阳生叶光合色素及光合作用对光环境的变化对的响应(Bertamini et al, 2006)、高山光环境对云杉生长和天然更新的影响(Diaci et al, 2005)、不同光环境条件对云杉表型可塑性(Silim et al., 2001;Lo et al., 2001;Niinemets et al., 2003;Kami et al., 2004)和生物量变化(Kennedy et al., 2007)的影响等。随着全球气候变暖,气候因子影响云杉幼苗生长、更新及其生理生态特性的研究不断增多(Yin et al., 2008)。不同海拔梯度上的云杉种群形态及生理特性上的变化实际上是其对不同环境因子变化的反应(Luo et al, 2005),而不同地区的云杉对营养元素的吸收能力在一定程度上反应出其生态适应性的大小(Major et al., 2007)。
天山云杉(Picea schrenkiana var. tianschanica)林分以天然纯林为主,是新疆分布最广、木材产量最高的珍贵针叶林,也是天山山地森林生态系统中最主要的植被类型,对天山的水源涵养、水土保持和林区其他生态系统的维持起着主导作用,构成天山乃至新疆森林生态系统的主体。有关天山云杉的研究多涉及种群结构及森林生产力等方面(王燕等, 2000;李明辉等, 2005),光合生理生态方面,李行斌等(2001)测定离体状态下、贾宏涛等(2005)自然状态下天山云杉针叶的光合速率,而对天然林分中天山云杉的基本光合特性还没有开展过研究。由于树木光合作用要受环境因子和树木自身光合特性的双重影响,特别是在自然生长环境中,处于不同生长阶段或年龄的树木,其光合特性的差异对生态系统物质循环与能量流动的贡献率是不同的(Cernusak et al., 2000;Kikuzawa et al., 2004;Mäkelä et al., 2004;Kolari et al., 2007)。比较自然生长环境中不同龄级天山云杉的光合生理生态特性,分析光合速率与环境因子之间的关系,有助于了解环境因子对自然生长的天山云杉影响。这将为进一步开展天山森林生态系统服务功能评价、天然林保护和合理经营奠定基础。
1 研究地点自然概况研究地点设在位于天山中部的国家林业局天山森林生态系统定位站(43°09′—43°28′ N,87°12′—87°50′ E)。该站位于新疆乌鲁木齐市南部乌鲁木齐县水西沟乡,属于天山北麓中部,占地面积369 hm2,其中林业用地266 hm2,有林地面积139 hm2。海拔变幅在1 908~2 710 m。年平均气温5 ℃, 极端最低气温-38 ℃,最高气温30 ℃;年降水量600~800 mm,无霜期150~160 d,属温带大陆性气候。土壤为山地灰褐色森林土。平均林龄为60~80年,最大年龄达150年。天山中部天山云杉林下代表性植物种有:宽刺蔷薇(Rosa platyacantha)、金丝桃叶绣线菊(Spiraea hypericifolia)、天山花楸(Sorbus tianschanica)、苔草(Carex spp.)、西伯利亚铁线莲(Clematis sibirica)、蓍草(Achillea millefolium)、高山羊角芹(Aegopodium podagraria)、天山羽衣草(Alchemila tianschanica)、珠芽蓼(Polygonum viviparum)、白花三叶草(Trifolium repens)、林地早熟禾(Poa nemoralis)、兰花菊(Cicerbita azurea)等。
天山森林生态定位站观测场对天山云杉的群落小气候、群落环境因子等进行着长期的定位观测,为将光合生理生态特性和环境因子等建立良好的匹配关系,本研究的林木测定地段设置在定位观测场所处的山坡(43°25′58″ N;87°28′20.4″ E)的林分内,海拔1 956 m,平均坡度为39°,坡向为32° NE。此山坡上的林分为天然次生的天山云杉纯林,林分的平均胸径17.3 cm,平均树高13.8 m。林下植被主要有小檗(Berberis)、蓍草、白花三叶草等,平均盖度约为35%。在此山坡的天山云杉林分中设置20 m×20 m的标准地3块(上、中、下坡各1块)进行每木检尺调查,根据标准地调查材料,按20年一个龄级,将所选取的林分分为4个龄级,即Ⅰ龄级(0~20年)、Ⅱ龄级(20~40年)、Ⅲ龄级(40~60年)及Ⅳ龄级(≥60年以上),绝大部分的天山云杉林木个体都分布在前3个龄级内,后1个龄级的个体分布数量较少。
2 材料与方法 2.1 研究材料的选取在研究地天然林分内,选择4个龄级中每个龄级接近年龄中值(即近10,30,50和70年生,在文中,我们分别用这些组中值代表 4个不同的龄级)的天山云杉各2株为代表,分别对每株当年生、1年生和2年生枝条的阳生叶和阴生叶进行光合生理生态指标测定,其中,Ⅰ、Ⅱ龄级测定部位选择在树体的中部,Ⅲ、Ⅳ龄级借助于搭梯将测定部位选择在2.2~2.5 m之间生长良好的枝条上。
2.2 试验方法 2.2.1 光合速率的测定于2005年的8—9月份间的晴天,使用LI-6400便携式光合作用测定系统完成天山云杉各项光合生理指标的测定。光合速率(Pn)的测定以小枝为基本测定单位进行,选择生长正常,针叶颜色、枝条长短、大小适中的枝条作为样枝,以保证测定的代表性。在同一枝条上按照明显的生长节确定当年生、1年生和2年生的生长段。测定时,在树体的阳面和阴面的同一层面上选择3根枝条作为重复,并标记重复序号。测定时间8: 00—20: 00时,时间间隔根据“早-中-晚”的顺序采用“2 h-1 h-2 h”的梯度进行,测定结果取平均值。同时测定的环境因子有光合有效辐射(PAR)、温度、相对湿度等。
2.2.2 光响应曲线的测定在晴天的光照最强的时段13:00—14:00之间,采用不同厚度白纱布遮挡的方法,用LI-6400-05簇状叶室测定不同龄级天山云杉阳生叶的光合速率随光照强度的变化曲线。首先用3~5层纱布(目的是达到根据光合日变化确定最大光合速率对应的光照强度)完全覆盖叶室约10 min,使叶片气孔完全打开;然后让枝条在全光照条件下使叶片气孔全部开放,测定其在最大光照强度下的净光合速率值,之后逐渐增加纱布的厚度达到控制光照强度的大小,完成光响应曲线的测定,测定设置3次重复。将实测的不同部位和不同叶龄天山云杉针叶净光合速率(Pn)对光合有效辐射(PPFD)的关系,用最小二乘法采用下述的经验方程进行拟合(Bassman et al., 1991):
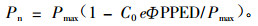
|
(1) |
式中,Φ为表观量子效率,C0为度量弱光下净光合速率趋于0的指标,通过适合性检验,拟和效果良好。并确定光补偿点和光饱和点。测定时的温度为(20.35±0.84) ℃,空气中CO2浓度约为(345.06±3.28) μmol·m-2s-1,相对湿度为38%±3.8%。
由于LI-6400-05簇状叶室没有人工光源,不能在全光照下使阴生叶的气孔完全开放,故阴生叶的光曲线未测定。
2.2.3 天山云杉针叶叶面积的确定不同年龄天山云杉叶面积采用排水法测定(周平等, 2002),计算公式

|
(2) |
式中,A为叶面积;V为天山云杉针叶体积(用排水法测定);L为针叶长度。计算出不同叶面积下的各项光合生理指标值。
2.2.4 光合速率与环境因子的相关性分析用SPSS 11.5分析以光合速率Pn为因变量,光照强度(PAR)、空气温度(Tair)、叶片温度(Tleaf)、空气CO2浓度(CO2-R)、叶室CO2浓度(CO2-S)、空气H2O浓度(H2O-R)、叶室H2O浓度(H2O-S)、空气相对湿度(RH-R)和叶室相对湿度(RH-S)为自变量的各因子间的相关性。
3 结果与分析 3.1 环境因子日变化研究区温度和光强的日变化见图 1。1天中的8:00—20:00时,环境和叶片温度的变化比较一致,叶片温度在12:00时达到最大,其值为(25.44±2.35) ℃,而环境温度在13: 00时也达到其最高峰值(26.23±2.52) ℃;早晚的温差变化不大,20: 00时的温度略低于8: 00时。光照强度的变化较大,从10: 00—14: 00时,光强维持较高水平,在14: 00时达到最大值(1707.94±87.39) μmol·m-2s-1。10: 00时之前和14: 00之后光照强度的上升和下降均很迅速。

|
图 1 温度和光照强度日变化 Figure 1 Diurnal changes of temperature and light intensity |
由图 2可知:环境中CO2浓度和H2O浓度在8: 00—20: 00时基本保持相对稳定的状态;CO2浓度在8: 00时和20: 00时的值略高于其他时间点的,而H2O浓度在这2个时间点的值略低于其他时间点的。叶室中的CO2浓度和H2O浓度在各个时间点的变化较大,H2O浓度随着时间的变化出现先增加后降低的趋势,CO2浓度的变化恰好相反,随着时间的变化呈现先降低后升高的趋势。早晚CO2浓度值是20: 00时的高于8: 00时的;而H2O浓度早晚的差异与此相反,为20: 00时H2O浓度低于8: 00时的值。叶室中H2O浓度值在12: 00时达到最大,为(15.09±1.68) μmolH2O·mol-1,CO2浓度在13: 00时达到最低值(351.89±4.78)μmolCO2·mol-1。

|
图 2 CO2和H2O浓度的日变化 Figure 2 Diurnal changes of CO2 and H2O concentration |
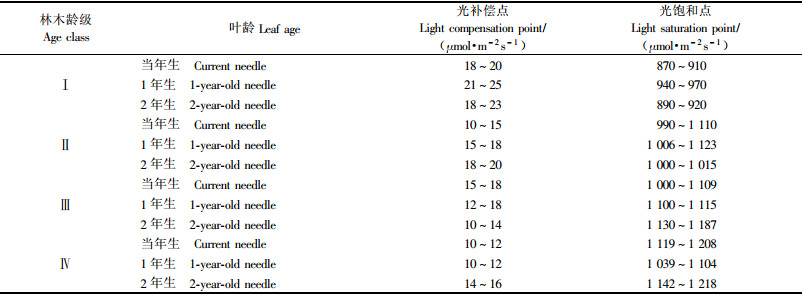
不同龄级天山云杉之间的光补偿点和光饱和点存在差异。光补偿点的变动范围在10~25 μmol·m-2s-1之间,光饱和点的变动范围在870~1 220 μmol·m-2s-1之间。随着年龄的增长,光补偿点有所下降,光饱和点有所增加。Ⅰ龄级天山云杉阳生叶的光补偿点明显高于其他年龄阶段,而光饱和点却低于其他的。其他年龄阶段的光补偿点和光饱和点比较接近,但Ⅳ龄级的光补偿点相对较低,光饱和点相对高于其他的。相同年龄林木上不同叶龄的光补偿点和光饱和点之间都有大小不一的差异;相同年龄林木、相同叶龄间的光补偿点和光饱和点的变化趋势一致(见表 1)。
|
|
最大光合速率(Pmax)通常代表植物的光合能力,在一定程度上决定着植物生长的快慢。比较不同年龄天山云杉最大净光合速率可发现,不同年龄以及相同年龄不同叶龄间的值也存在差异(图 3)。各年龄阳生叶的最大净光合速率均明显大于阴生叶的最大净光合速率。

|
图 3 不同年龄天山云杉最大净光合速率 Figure 3 The maximum net photosynthetic rate of the spruce in different age classes P.schrenkiana var. tianschanica A:Ⅰ龄级Age class Ⅰ; B: Ⅱ龄级Age class Ⅱ C:Ⅲ龄级Age class Ⅲ; D: Ⅳ龄级Age class Ⅳ |
阳生叶中,4个龄级中相同叶龄的阳生叶最大净光合速率随着龄级的增加表现出先增加后有所降低的趋势,而且当年生叶的最大净光合速率值要高于1年生和2年生叶的值(Ⅱ龄级1年生叶除外)。其中,当年阳生叶净光合速率大小顺序为Ⅲ龄级>Ⅳ龄级>Ⅰ龄级>Ⅱ龄级,其最大净光合速率值分别为(17.51±0.41),(16.36±0.05),(1.94±0.97)和(0.77±0.15) μmol CO2·m-2s-1,但Ⅰ到Ⅱ龄级的净光合速率增长比较小,从Ⅱ到Ⅲ龄级的增长迅速且达到最大光合能力,到Ⅳ龄级时的净光合速率值下降。按照上述林木龄级排序,1年和2年阳生叶净光合速率的大小顺序为Ⅲ龄级>Ⅳ龄级>Ⅱ龄级>Ⅰ龄级,其最大净光合速率值分别为(14.63±1.51),(13.6±1.21),(13.12±1.90),(9.74±0.30)和(15.64±0.07),(9.97±0.12),(8.09±0.36),(6.89±0.18) μmol CO2·m-2s-1。3个叶龄的天山云杉中,其Ⅰ和Ⅱ龄级,Ⅲ和Ⅳ龄级阳生针叶最大净光合速率之间差异均不显著(P>0.05)。Ⅰ龄级天山云杉3个叶龄最大净光合速率间差异显著(P<0.05),Ⅱ、Ⅲ、Ⅳ龄级之2年生阳生叶分别与其他2个叶龄间的差异显著(P<0.05)。从Ⅲ到Ⅳ龄级当年生阳生叶的净光合速率减少了6.56%,1年生叶的减少了15.93%,2年生的降低幅度最大,减少了36.25%。可见,随着天山云杉林木年龄的增加,不同年龄阳生叶片的光合速率都会降低。
阴生叶的最大净光合速率随着云杉年龄的增加其值是下降的,而且各个年龄中均为当年生阴生叶的最大净光合速率分别大于1年生和2年生的,即阴生叶的光合能力随着林木年龄和叶龄的增加而降低的。Ⅰ到Ⅳ龄级天山云杉当年生阴生叶的最大净光合速率值分别为(5.42±0.11),(2.44±0.08),(1.94±0.97),(0.77±0.15) μmolCO2·m-2s-1;1年生阴生叶的最大净光合速率分别为(4.43±0.04),(1.88±0.001),(0.37±0.025),(0.61±0.004) μmolCO2·m-2s-1;2年生阴生叶的最大净光合速率分别为(2.69±1.19),(1.98±0.29),(0.44±0.12),(0.32±0.04) μmolCO2·m-2s-1。但4个龄级不同叶龄之间阴生叶的差异均不显著(P>0.05)。
3.3.2 不同年龄天山云杉净光合速率日变化不同年龄天山云杉阳生叶净光合速率的日变化均为单峰曲线(图 4A)。在10: 00—16: 00时,各年龄天山云杉阳生叶的净光合速率值均明显高于其他时间点,且保持较高的值。在8: 00时,净光合速率均很低,随着光照强度和温度的升高,净光合速率在10: 00时迅速升高,甚至达到最大值,部分叶龄的叶片在12: 00时或13: 00时达到最大;16: 00时之后的光合速率值随着光照强度的明显降低而迅速下降,在20: 00时部分阳生针叶开始呼吸作用。对比4个龄级天山云杉阳生叶的光合速率值可知,当年生阳生叶的光合速率值要高于1年生和2年生的值。其中Ⅲ龄级林木的光合速率值要高于其他的,而且此年龄级不同叶龄的光合速率值比较接近。其中,Ⅰ和Ⅲ龄级林木的当年生阳生叶净光合速率在10: 00时达到最大值,Ⅱ和Ⅳ龄级林木的分别在12: 00时和13: 00时达到最大,但Ⅰ和Ⅱ龄级林木的最大光合速率值低于Ⅲ和Ⅳ龄级林木的。Ⅲ龄级林木的当年生阳生叶净光合速率从10: 00—16: 00时段均保持较高的水平,而其他年龄林木的当年生阳生叶在10: 00—14: 00时段净光合速率的值高于其他时段;Ⅰ和Ⅱ龄级林木的1年生阳生叶的净光合速率在10: 00时达到最大值,Ⅲ和Ⅳ龄级林木的在12: 00时达到最大,且在10: 00—16: 00时段内均保持较高的净光合速率值。4个龄级2年生阳生叶的净光合速率均在12: 00时达到最大值。

|
图 4 不同年龄天山云杉的净光合速率日变化 Figure 4 Dinurnal change of net photosynthetic rate of P. schrenkiana var tianschanica in different age classes A:阳生叶Sun-growing leaf; B:阴生叶Shade-growing leaf |
4个龄级天山云杉阴生叶净光合速率的日变化趋势不同,Ⅰ龄级林木为单峰曲线,其他年龄林木表现出不很明显的双峰曲线(图 4B)。在光合高峰期,Ⅰ龄级天山云杉的净光合速率值高于其他年龄林木的,其不同叶龄的净光合速率随着温度和光照强度的变化出现先升高后降低的日变化趋势;其中,当年生和2年生阴生叶在13: 00时净光合速率值最大,1年生阴生叶在12: 00时净光合速率值达到最大。Ⅱ龄级林木当年生和1年生阴生叶净光合速率在10: 00时达到第1个高峰后又降低,在13: 00时为第1个谷底,在14: 00时又上升到第2个高峰,之后又降低;2年生阴生叶从8: 00—13: 00时净光合速率一直上升,14: 00时下降谷底后在15: 00时又达到第2个峰值,之后又开始下降。在20: 00时,各叶龄阴生叶净光合速率值的大小顺序为当年生>1年生>2年生。Ⅲ和Ⅳ龄级的天山云杉不同叶龄的净光合速率在8: 00时的值相对较高,之后开始下降,表现出呼吸速率增加的趋势,在12: 00时和10: 00时分别达到最大的呼吸速率值,之后呼吸速率再降低,到16: 00时达到第2个光合高峰。在20: 00时,1年生的净光合速率均大于当年生和2年生的值。
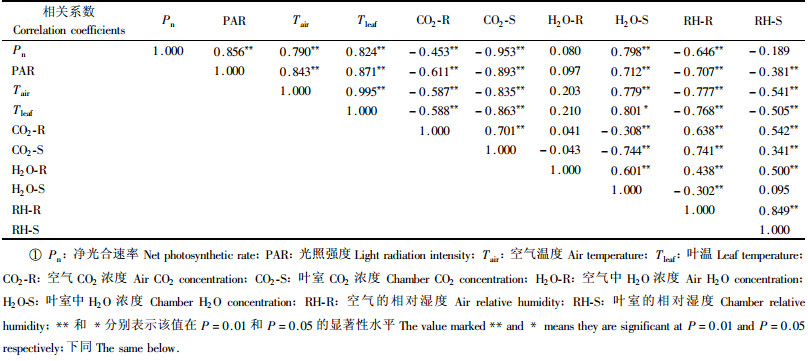
3.4 天山云杉光合速率与环境因子的相关性本研究选择和净光合速率同步测定的9个环境因子为自变量,与其进行相关分析(表 3~4)。从表 3中看出,阳生叶净光合速率(Pn)与PAR、Tair、Tleaf、CO2-R、CO2-S、H2O-S和RH-R之间具有显著的相关性。其中,与PAR的正相关性最大(R=0.856);其次为Tleaf(R=0.824),其余的依次为H2O-S(R=0.798)、Tair(R=0.790);与CO2-S和的负相关性最大(R=-0.953),这可能是因为密闭叶室内云杉针叶的光合作用要消耗一定量的O2,同时进行的光呼吸要释放一定量的CO2,这2种生理过程会影响叶片周围CO2浓度的变化,适宜的CO2浓度会促进光合作用的进行,而高浓度的CO2会抑制叶片光合作用;其次为RH-R (R=-0.646)和CO2-R (R=-0.453)。Pn与H2O-R和RH-S之间无相关性。
|
|
|
|

从表 4中可以看出,阴生叶的净光合速率(Pn)只与PAR之间具有极显著的正相关性(R=0.782 5),与RH-R之间具有极显著的负相关性(R=-0.220 4)。与其他环境因子之间无相关性。空气温度与湿度之间具有极显著的相关性。
4 讨论天山云杉是天山森林生态系统的主体,树木光合作用是森林生态系统能量流动和物质循环的基础,是决定森林生产力最重要的生理过程,选择自然生长状态下天山云杉进行研究,既消除实验室条件下人为因素的干扰,使实验结果更现实化,也在一定尺度上为天山云杉群落及其森林生态系统碳循环研究提供基础数据。但光合作用要受外界各种环境因素和树木自身生物学特性(如:针叶结构)的双重影响。这为研究天然林分中天山云杉的光合特性增加难度,使得研究结果带有一定的不确定性。但作为自然生态系统的主体,由于天山山脉气候、环境的特殊性,加之生态系统的脆弱性,急需对该生态系统主体——天山云杉天然林分进行生理生态特性的基础研究,以便为开展该生态系统其他服务功能的研究奠定基础。
1) 光响应曲线是理解光能驱动下光合有效辐射和净光合速率间关系的基础。它所指示叶片的光饱和点与光补偿点反映植物对光照条件的要求,分别体现对强光和弱光的利用能力(Ye, 2007)。天山云杉不同叶龄的光补偿点和光饱和点在数值上存在一定的差别,但均体现出较低的光补偿点和较高光饱和点。不同种类以及相同种类不同地区的云杉的光补偿点存在差异,天山云杉对光环境的适应性较强。本文测定的不同叶龄天山云杉的光补偿点在10~25 μmol·m-2s-1范围之间,低于贾宏涛等(2005)的测定天山云杉光补偿点的值88.33 μmol·m-2s-1;也低于红皮云杉(Picea koraiensis)的光补偿点值47 μmol·m-2s-1(张放等,2003)以及林芝云杉(P. likiangensis var. linzhiensis)的值107.5 μmol·m-2s-1(肖文发等,2003)。本研究得出天山云杉光饱和点范围在870~1 220 μmol·m-2s-1之间,与贾宏涛等(2005)提出的天山云杉的光饱和点应高于2 000 μmol·m-2s-1的研究结果存在明显差异。本试验在9月份进行,晴天的最大光强可以达到1 800 μmol·m-2s-1以上,但该光照条件下的净光合速率不能达到最大。由于新疆地处我国温带,森林属于亚高山森林,而高山森林的西藏林芝云杉在晴天的光饱和点仅有770 μmol·m-2s-1(肖文发等,2003)。
2) 在不同的年龄阶段,树木光合能力的大小也是有差别的,根据本文的研究可以将天山云杉在整个生长周期中同化CO2的能力大致可以分为3个阶段:一是Ⅰ、Ⅱ龄级到Ⅲ龄级阶段的光合速率缓慢增加阶段;二是Ⅲ龄级到Ⅳ龄级光合速率迅速增加,并达到最大同化能力的阶段;三是在Ⅳ龄级后期到成年以后,随着年龄的增加,树木光合产物中有较大比例是用来维持自身体内的各种代谢过程,在数值的大小上体现光合速率值在降低的阶段。由于受环境因子的影响,各个生长阶段林木的净光合速率日变化也是有差异的。本文研究的天山云杉阳生叶净光合速率的日变化为单峰曲线,而且峰值出现在10: 00时左右,而且随着林木树龄的不同,峰值的大小也存在差异,其大小顺序为Ⅲ龄级>Ⅳ龄级>Ⅱ龄级>Ⅰ龄级。青海云杉(Picea crassifoia)(汪杰等,2004)和黑云杉(P.mariana)(杨丽涛等,2000)Pn的日变化也为典型的单峰曲线,与贾宏涛等(2005)的结果不一致。
Ⅰ龄级林木阴生叶的净光合速率日变化为单峰曲线,峰值出现在13: 00时左右。这是由于这些树体相对其他年龄的树体来说较矮小,随着时间的变化,感受的光照强度较大,净光合速率值基本为正值,即1天中的大部分时间均在进行有机物的积累。但是Ⅱ龄级、Ⅲ龄级和Ⅳ龄级林木阴生叶的净光合速率却表现出与上述不同的双峰曲线,峰值分别出现在8: 00时和16: 00时,比阳生叶净光合速率高峰出现时间略有滞后,即阴生叶在日变化过程中提前出现“午休现象”,而引起“午休”的因素很多,最主要的是温度、光照和水分。由于测定时叶片所处的温度在20~25 ℃之间,并不能引起酶的钝化和叶绿体和细胞质结构的破坏,因此由温度引起光合“午休”的可能性很小。对阴生叶净光合速率与环境因子的相关分析结果说明:导致这种现象的主要原因来源于光照强度和水分(或湿度),这与Talbott等(2003)的研究结果具有一致性。Bertamini等(2006)的研究表明:挪威云杉(Picea abies)阴生叶的叶绿素、类胡萝卜素含量及RuBP羧化酶活性明显低于阳生叶的。但从光照强度来分析,由于Ⅱ龄级、Ⅲ龄级和Ⅳ龄级的树冠相对Ⅰ龄级的大,虽然实验取样过程中尽量减少了周围环境的异质性对树木光合作用的影响,但高大树冠内不同枝条间对光照的遮蔽也会成为影响其光合作用过程不可避免的因子;而较大龄级树木的阴生叶在长期的适应过程中,对光照的敏感远高于温度,弱的光照可能会导致光抑制。在测定时段内,研究区降水较少,较低的湿度也成为抑制光合产物积累的因素之一。
3) 不同年龄的天山云杉,不论是阳生叶还是阴生叶,其净光合速率均与光照强度(PAR)成极显著的正相关性。与空气相对湿度成极显著的负相关。阳生叶的净光合速率与CO2浓度成极显著的负相关,而阴生叶与CO2浓度之间无相关性。关于CO2倍增可以提高作物光合速率的试验,在国内外开展了诸多相关的研究。而这种促进作用随不同季节而变化(蒋高明,1995)。Marek等(1997)研究表明不同生长期挪威云杉光合速率对CO2浓度梯度的响应也存在时间性。本研究在自然环境中CO2浓度与光合速率负相关的结果与上述观点具有一致性。
贾宏涛, 赵成义, 蒋平安, 等. 2005. 天山云杉林地生态系统CO2源/汇关系研究. 干旱区地理, 28(2): 244-247. DOI:10.3321/j.issn:1000-6060.2005.02.021 |
蒋高明. 1995. 全球大气二氧化碳浓度升高对植物的影响. 植物学通报, 12(4): 1-7. |
李明辉, 何风华, 刘云, 等. 2005. 天山云杉种群空间格局与动态. 生态学报, 25(5): 1000-1006. DOI:10.3321/j.issn:1000-0933.2005.05.009 |
李行斌, 白志强, 郭仲军, 等. 2001. 天山云杉等主要树种光合作用初探. 新疆农业科学, 38(2): 62-65. DOI:10.3969/j.issn.1001-4330.2001.02.004 |
汪杰, 张盹明. 2004. 两种针叶树种光合特性研究. 水土保持学报, 18(5): 159-162. DOI:10.3321/j.issn:1009-2242.2004.05.039 |
王燕, 赵士洞. 2000. 天山云杉林生物生产力的地理分布. 植物生态学报, 24(2): 186-190. DOI:10.3321/j.issn:1005-264X.2000.02.011 |
肖文发, 韩景军, 郭志华, 等. 2003. 西藏林芝云杉针叶净光合速率对环境因子的响应. 林业科学研究, 16(3): 299-305. DOI:10.3321/j.issn:1001-1498.2003.03.009 |
杨丽涛, ArkebauerT J, Walter-SheaEA. 2000. Diurnal variation in gas exchange of aspen, hazelnue, jack pine and black spruce. 植物生态学报, 24(4): 408-419. DOI:10.3321/j.issn:1005-264X.2000.04.005 |
张放, 胡万良, 孔祥文. 2003. 红皮云杉幼树不同光环境下生理生态特征研究. 辽宁林业科技, 5: 1-4. |
周平, 李吉跃, 招礼军. 2002. 北方主要造林树种苗木蒸腾耗水特性研究. 北京林业大学学报, 24(5/6): 50-55. |
Bassman J, Zwier J C. 1991. Gas exchange characteristics of Populus trichocarpa, Populus deltoids and Populus trichocarpa ×P. deltoids clone. Tree Physiol, 8: 145-159. DOI:10.1093/treephys/8.2.145 |
Becker P. 2000. Competition in the regeneration niche between conifers and angiosperms: Bond's slow seedling hypothesis. Functional Ecology, 14: 401-412. DOI:10.1046/j.1365-2435.2000.00455.x |
Bertamini M, Muthuchelian K, Nedunchezhian N. 2006. Shade effect alters leaf pigments and photosynthetic responses in Norway spruce (Picea abies L.) grown under field conditions. Photosynthetica, 44(2): 227-234. DOI:10.1007/s11099-006-0011-z |
Cernusak L A, Marshall J D. 2000. Photosynthetic refixation in branches of Western White Pine. Functional ecology, 14: 300-311. DOI:10.1046/j.1365-2435.2000.00436.x |
de Chantal M, Leinonen K, Kuuluvainen T, et al. 2003. Early response of Pinus sylvestris and Picea abies seedlings to an experimental canopy gap in a boreal spruce forest. For Ecol Manage, 176: 321-336. DOI:10.1016/S0378-1127(02)00273-6 |
Diaci J, Pisek R, Boncina A. 2005. Regeneration in experimental gaps of subalpine Picea abies forest in the Slovenian Alps. Eur J Forest Res., 124: 29-36. DOI:10.1007/s10342-005-0057-7 |
Kamiska-Roek E, Pukacki P M. 2004. Effect of water deficit on oxidative stress and degradation of cell membranes in needles of Norway spruce (Picea abies). Acta Physiologiae Plantarum, 26(4): 431-442. DOI:10.1007/s11738-004-0034-7 |
Kennedy S, Black K, O'Reilly C, et al. 2007. The impact of shade on morphology, growth and biomass allocation in Picea sitchensis, Larix×eurolepis and Thuja plicata. New Forests, 33: 139-153. DOI:10.1007/s11056-006-9019-2 |
Kikuzawa K, Shirrakawa H, Suzuki M, et al. 2004. Mean labor time of a leaf. Ecological Research, 19: 365-374. DOI:10.1111/ere.2004.19.issue-4 |
Kolari P, Lappalainen H K, Hänninen H, et al. 2007. Relationship between temperature and the seasonal course of photosynthesis in Scots pine at northern timberline and in southern boreal zone. Tellus B, 59(3): 542-552. DOI:10.1111/j.1600-0889.2007.00262.x |
Lo E, Wang Z M, Lechowicz M J, et al. 2001. Adaptation of the LIGNUM model for simulations of growth and light response in jack pine. For Ecol Manage, 150: 279-291. DOI:10.1016/S0378-1127(00)00587-9 |
Luo J X, Zang R G, Li C Y. 2005. Physiological and morphological variations of Picea asperata populations originating from different altitudes in the mountains of southwestern China. For Ecol Manage, 221: 285-290. |
Lusk C H, Wright I, Reich P B. 2003. Photosynthetic differences contribute to competitive advantage of evergreen angiosperm trees over evergreen conifers in productive habitats. New phytologist, 160: 329-336. DOI:10.1046/j.1469-8137.2003.00879.x |
Major J E, Mosseler A, Barsi D C, et al. 2007. Comparative nutrient economy, stable isotopes, and related adaptive traits in Picea rubens, Picea mariana, and their hybrids. Tree, 21: 677-692. DOI:10.1007/s00468-007-0161-z |
Mäkelä A, Hari P, Berninger F, et al. 2004. Acclimation of photosynthetic capacity in Scots pine to the annual cycle temperature. Tree Physiol, 24: 369-378. DOI:10.1093/treephys/24.4.369 |
Marek M V, Šprtová M, Kalina J. 1997. The photosynthetic irradiance-response of Norway spruce exposed to a long elevation of CO2 concentration. Photosynthetica, 33(2): 259-268. DOI:10.1023/A:1022120531807 |
Nedbal L, Červený J, Rascher U, et al. 2007. E-photosynthesis: a comprehensive modeling approach to understand chlorophyll fluorescence transients and other complex dynamic features of photosynthesis in fluctuating light. Photosynth Res, 93: 223-234. DOI:10.1007/s11120-007-9178-9 |
Niinemets Ü, Lukjanova A. 2003. Needle longevity, shoot growth and branching frequency in relation to site fertility and withincanopy light conditions in Pinus sylvestris. Ann For Sci, 60: 195-208. DOI:10.1051/forest:2003012 |
Niinemets Ü. 2007. Photosynthesis and resource distribution through plant canopies. Plant, Cell and Envinonment, 30: 1052-1071. DOI:10.1111/pce.2007.30.issue-9 |
Portsmuth A, Niinemets Ü. 2007. Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Functional Ecology, 21(1): 61-77. |
Silim S N, Guy R D, Patterson T B, et al. 2001. Plasticity in water-use efficiency of Picea sitchensis, P. glauca and their natural hybrids. Oecologia, 128: 317-325. DOI:10.1007/s004420100659 |
Stenberg P, Kangas T, Smolander H, et al. 1999. Shoot structure, canopy openness, and light interception in Norway spruce. Plant, Cell & Environment, 22(9): 1133-1142. |
Talbott L D, Rahveh E, Zeiger E. 2003. Relative humidity is a key factor in the acclimation of the stomatal response to CO2. Journal of Experimental Botany, 54(390): 2141-2147. DOI:10.1093/jxb/erg215 |
Wilson K B, Baldocchi D D, Hanson P J. 2001. Leaf age affects the seasonal pattern of photosynthetic capacity and net ecosystem exchange of carbon in a deciduous forest. Plant, Cell and Environment, 24: 571-583. DOI:10.1046/j.0016-8025.2001.00706.x |
Ye Z P. 2007. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa. Photosynthetica, 45(4): 637-640. DOI:10.1007/s11099-007-0110-5 |
Yin H J, Liu Q, Lai T. 2008. Warming effects on growth and physiology in the seedings of the two conifers Picea asperata and Abies faxoniana under two contrasting light conditions. Ecol Res, 23: 459-469. DOI:10.1007/s11284-007-0404-x |