 2009, Vol. 45
2009, Vol. 45文章信息
- 王有年, 邢彦峰, 周士龙, 孟海玲, 关伟, 师光禄.
- Wang Younian, Xing Yanfeng, Zhou Shilong, Meng Hailing, Guan Wei, Shi Guanglu
- 观光桃园昆虫群落结构与优势种生态位
- Structure, Dynamics and Niche of Dominant Population of Insect Community in Peach Orchards of Beijing
- 林业科学, 2009, 45(4): 88-94.
- Scientia Silvae Sinicae, 2009, 45(4): 88-94.
-
文章历史
- 收稿日期:2007-11-29
-
作者相关文章



2. 农业部都市农业(北方)重点开放实验室 北京 102206;
3. 北京市平谷区果品办公室 北京 101200
2. Key Laboratory of Urban Agriculture (North) of Ministry of Agriculture Beijing 102206;
3. Fruit Agency of Pinggu District of Beijing Beijing 101200
桃树原产于我国,是北京乃至华北地区的重要果树之一。近年来随着旅游产业的不断发展,北京旅游观光采摘桃园的面积逐年扩大,并具有一定的规模,为提高果农的经济收入起到了积极的作用。然而频繁发生的病虫害,直接影响着桃的产量、品质和经济效益,这就要求我们运用有害生物综合治理的原理与方法,改进桃园病虫害的管理水平。桃园生态系统中栖息着多种生物类群,其中昆虫种类多、数量大、发生时间长, 是桃园生态系统的重要生物类群, 对其群落的研究是探讨桃园害虫结构动态、进行种群数量预测与优化调控的主要策略。昆虫群落的时序结构与动态的变化,直接反映着昆虫与植物互作的生态效应,对其系统研究有利于昆虫群落多样性与稳定性的探究,正确评价有害生物生态调控的效果。昆虫生态位直接反映其在环境中的地位和其他生物间的相互关系,有关方面的研究已有较多的报道(黄保宏等,2005;师光禄等, 2005;2006a;2006b;邹运鼎等,2003;李昌根等, 2005;周洪旭等, 2004; 万方浩等,1986),但在群落的水平上系统调查分析桃园昆虫群落结构与优势种,并根据桃园整个昆虫群落的变化规律制定主要害虫综合治理对策的报道甚少。本文系统地调查了北京地区桃园主要害虫及天敌群落的结构时序动态与生态位,研究了害虫与天敌之间的相互关系,旨在为桃园害虫的可持续控制提供科学依据。
1 试验区概况与研究方法 1.1 试验区概况试验区设在北京市平谷区大华山镇苏子峪村,海拔约400 m,40°10′—40°26′N,116°54′—117°06′E,属于燕山山脉的半干旱山区,该村东西长约2.9 km,南北长约3.0 km,年平均气温8~10℃,无霜期约180~190天,年均降水量400~600 mm,主要集中在6—9月。土壤属温暖带半湿润地区的褐土地带,有机质含量1.7 g·kg-1,全氮含量0.71 g·kg-1,速效磷22.77 μg·g-1,土壤pH值4.5~7.5,土壤偏弱酸性。主要以生产蜜枣(Ziziphus jujube)、大桃(Prunus persica)、核桃(Juglans regia)、柿子(Diospyros kaki)、蜜梨(Pyrus pyrifolia)和板栗(Castanea mollissima)等果品为主。全村桃树面积60 hm2,约占果树面积的32%,品种主要为久保和蟠桃系列,树龄6年,树高2~2.5 m,桃园按相关部门制定的有机果品标准化生产的技术措施进行管理。
1.2 调查方法调查于2006年4—10月进行,采用棋盘式5点取样,每样点选取具有代表性的桃树2株,每株树冠分为上、中、下3个部位,并将各部位分为东、西、南、北4个方位,每株树共调查12个点,每位点选择2枝2~3年生(大于15 cm)枝条,每10天调查1次,采用网捕与目测相结合的方法,观测和记录此范围内昆虫(包括蜘蛛和螨类)的种类与数量,暂不能定名的物种,用盛有75%酒精的佩尼西林小瓶杀死保存,统一编号后带回室内鉴定其目、科和种(黄保宏等,2005)。
1.3 分析方法群落结构的测定以月为单位,计算各月内同种昆虫数量的总和,然后进行多样性、均匀度、优势度的分析。用Shannon-Wiener平均信息量指数H′分析群落多样性: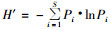
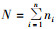

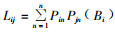

经系统调查,共收集昆虫34种,隶属9目25科,其中植食性种类18种,天敌昆虫11种,中性昆虫5种。鳞翅目、同翅目、半翅目和鞘翅目为该群落的主要类群,这4个目的种类占全部种类的91.2%,个体数占全部样本量的95.6%。桃蛀果蛾(Carposina niponensis)、梨小食心虫(Pseudaulacaspis pentagona)、桃蛀野螟(Dichocrocis punctiferalis)、桃潜蛾(Lyonetia clerkella)、桑白盾蚧(Pseudaulacaspis pentagona)、桃蚜(Myzus persicae)、小花蝽(Orius sauteri)、异色瓢虫(Leis axyridis)、黑带食蚜蝇(Epistrophe balteata)、叶色草蛉(Chrysopa phyllochroma)、黑缘红瓢虫(Chilocorus rubidus)等是桃园昆虫优势种。
数据处理分析结果见图 1。由图可见,桃树萌动露绿前,昆虫群落结构较为简单,大多处于休眠状态。开花展叶前,植食类昆虫开始活动,并寻找适当场所开始取食或产卵,此时种群数量开始上升,物种数与个体数明显增加,尤其幼芽与花器的害虫较多,到4月中旬后,桑白盾蚧、梨小食心虫等害虫为此阶段的优势种,桃树被害症状十分明显。进入5月份,昆虫种群数量明显增多,密度明显增大,尤其是桃蚜、蚧虫类以及叶螨类等刺吸食害虫为优势种。此时昆虫无论个体数与丰富度,还是多样性均高于4月份的值。与4月份相同的是害虫亚群落的丰富度与个体数均高于天敌亚群落,但多样性与均匀度则正好相反,不言而喻,天敌亚群落发展比较均衡稳定,利于桃园应用天敌调控害虫的发生。由此可见,在有机化管理的桃园中,桃树萌动露绿前,应该采取农业与生物防治措施,压低这些桃树害虫的发生基数,保护和提高天敌的种群数量, 是有机观光桃园害虫生态调控的重要措施之一,随着桃树的发育, 昆虫的群落结构也在发生变化。

|
图 1 桃园昆虫丰富度、多样性、均匀度和总个体数时序动态 Figure 1 Time dynamics of richness, diversity, evenness and individual number of insect community in peach orchard |
进入6—7月,由于桃园昆虫的食物丰富,气候适宜,食叶、食花、食果害虫同时出现,天敌种类也随之增多,形成复杂的食物网络结构,是全年群落多样性、丰富度、均匀度较高,群落自控能力较强、较稳定的时期。此期害虫亚群落与天敌亚群落的参数与其他不同发育阶段相比均较大。从图 1明显可见,昆虫的丰富度与多样性明显上升,而个体数开始下降,害虫亚群落无论从个体数与丰富度,还是多样性与均匀度均高于天敌亚群落,天敌亚群落的多样性变化一直比较平稳。说明此阶段有机观光桃园天敌亚群落的稳定性较好。4—7月昆虫群落的丰富度呈现上升趋势,波动的频率趋于平稳,6—7月害虫亚群落的多样性与均匀度指数呈大幅度上升趋势,但总的趋势是天敌亚群落的种群数量随害虫亚群落数量的增加而缓慢地提高,相对滞后于害虫亚群落的增长,但具有较强的跟随关系。可见此阶段应结合夏剪与保果等叶面施肥与相应的管理手段,进行调治病虫害是提高天敌亚群落的种群数量,降低害虫亚群落的种群密度,改善桃园昆虫生态环境的重要措施之一。到8—10月,天敌亚群落的种群数量及其多样性与均匀度高于害虫亚群落,但丰富度则低于害虫亚群落,说明该阶段天敌亚群落中有一定数量的优势种,对害虫亚群落的重要种类有一定的调控作用。因此,桃果采摘后,应该采取积极的措施,例如通过刮树皮、杜树洞、结合修剪、清理桃园枯枝落叶,以及深翻树盘等方法来压低越冬害虫的基数。
2.2 桃园主要害虫及天敌空间生态位分析 2.2.1 空间生态位分析将主要害虫及天敌空间生态位的计算结果列于表 1。由表 1可见,在空间格局上,害虫的生态位宽度依次为:桑白盾蚧(0.704 4)>桃蚜(0.670 6)>桃潜蛾(0.643 9)>梨小食心虫(0.632 5)>桃蛀野螟(0.596 2)>桃蛀果蛾(0.574 3),说明这些害虫在各资源的序列上均匀分布。由于桑白盾蚧是与刺吸式口器危害,它几乎分布于桃树的各个部位,因此生态位宽度相对较大,桃蚜与桃潜蛾主要危害寄主的叶部位,它的生态位宽度要小于桑白盾蚧,梨小食心虫主要危害桃树的顶梢,被害后的桃梢枯萎下垂,症状十分明显,极易发现,但桃蛀野螟与桃蛀果蛾主要以幼虫钻蛀果实,食心危害,因此生态位宽度相对较小。但这些害虫是目前桃园的主要害虫,应结合不同的桃园管理与环境条件加以适当的调控。天敌的生态位宽度依次为:黑缘红瓢虫(0.720 5)>异色瓢虫(0.713 1)>黑带食蚜蝇(0.681 7)>叶色草蛉(0.689 4)>小花蝽(0.673 2),可见他们在资源的利用上是均匀的。调查发现,黑缘红瓢虫和异色瓢虫主要捕食蚧虫类、蚜虫类害虫,而蚧虫与蚜虫是桃园的主要刺吸类害虫,其种群数量相对较大,危害较重的树常出现顶梢枯萎,早期落叶,果实萎缩,树势衰弱等明显症状,因此,对瓢虫等捕食类天敌的保护是调控桃园主要刺吸类害虫的有效措施之一。黑带食蚜蝇与叶色草蛉主要捕食蚜虫类害虫,它的生态位宽度显然要较小于黑缘红瓢虫和异色瓢虫,但在调控蚜虫的种群数量上有一定的作用。
|
|

桃蛀果蛾与桃蛀野螟、桃蚜与桃潜蛾在空间上具同域性,相互竞争空间资源,因此生态位重叠较大;黑缘红瓢虫和异色瓢虫两种捕食性昆虫与桑白盾蚧和桃蚜两种害虫间的生态位重叠均较大,桃蚜与黑带食蚜蝇的生态位重叠也较大,说明这些天敌具有相同的空间占有量,对这些害虫具有一定的调控作用。黑缘红瓢虫、异色瓢虫、黑带食蚜蝇三者之间的生态位重叠也较大,可见其种群数量多时会发生食物竞争,从而增强了对害虫亚群落的控制作用,也说明天敌的食性广泛。异色瓢虫和黑缘红瓢虫2种瓢虫与桑白盾蚧的生态位相似性比例指数较大,黑带食蚜蝇与桃蚜次之,其他种类也均较高,且它们之间的差距较小,说明这些害虫与天敌的种群数量都维持在比较高的水平,对桃园空间生态位的占据上相似程度较高。可见桃园采取正确的测报与调控措施,对保护天敌去控制害虫的发生危害具有十分重要的意义。
2.2.2 时间生态位分析时间虽然不是物种竞争的直接对象,但它是物种竞争或共存得以实现的必要条件。由表 2可见,害虫的时间生态位宽度依次为:桑白盾蚧(0.821 6) >桃蚜(0.818 4) >桃潜蛾(0.801 4) >梨小食心虫(0.745 0) >桃蛀野螟(0.580 4)>桃蛀果蛾(0.549 7)。从田间调查结果来看,除冬季外,桑白盾蚧周年均可发生危害桃树,世代重叠明显,除卵外,各虫态均可周年在桃树的各个部位取食,且虫口密度较大,为此时间生态位宽度也较大,受此虫危害严重的寄主常出现枝条枯萎,因而需采取措施予以控制。桃蚜从树叶流动后开始活动危害,以后逐渐扩散于叶背、嫩茎顶梢上吸食汁液,大量发生时,常密集于嫩梢、叶部作不规则卷曲梢上冒油,阻碍了树体生长,影响果实的产量与花芽的形成,大大削弱了树势。危害到10月间开始产卵越冬,不难理解,桃蚜在桃树上的危害几乎发生在全年,因而时间生态位相对较大。桃潜蛾是以蛹越冬,开始发生危害的时间相对较后,4月下旬可见到被害症状,但持续到桃树落叶均有被害状,并在被害叶上结白色小茧越冬,可见时间生态位宽度要小于桑白盾蚧与桃蚜。梨小食心虫是以幼虫钻蛀危害桃梢,尤其在桃梨混栽果园,其发生密度在春季较大,危害较重,除危害桃梢外,还可危害桃果,主要发生在6,7月间。8月及以后主要危害梨果,与前几种害虫比较,时间生态位宽度较小。桃蛀野螟与桃蛀果蛾均以幼虫蛀果危害,但两者的被害症状明显不同。桃蛀野螟是以幼虫从果梗基部沿果核蛀入果心,食害核仁与果肉,且果外蛀孔明显,周围对有大量红褐色虫粪,两果相接处危害较重。调查发现桃蛀野螟有1果产多卵的习性,且一果中常有数条幼虫寄生危害。桃蛀果蛾1果只产卵1粒,蛀孔较小,且一果中仅有1头幼虫寄生危害,桃蛀果蛾的幼虫则是多从萼凹或果面处蛀入, 并直达果核周围食害果肉,虫粪常留在果核周围。桃蛀野螟幼虫发生危害期比桃蛀果蛾提前1~2个月,因此桃蛀野螟的时间生态位宽度要相对大于桃蛀果蛾。
|
|
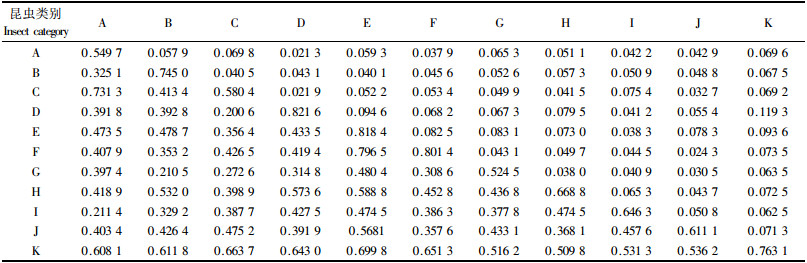
天敌的时间生态位宽度依次为:黑缘红瓢虫(0.763 1)>异色瓢虫(0.668 8)>黑带食蚜蝇(0.646 3)>叶色草蛉(0.611 1)>小花蝽(0.524 5)。黑缘红瓢虫与异色瓢虫是蚧虫与蚜虫的重要天敌,调查发现,桃园蚧虫与蚜虫是害虫亚群落中的优势种群,不仅虫口密度大,而且危害持续的时间也较长,黑缘红瓢虫与异色瓢虫的跟随效应明显,是蚧虫与蚜虫的主要捕食性天敌,可见它们的时间生态位宽度相对较大,黑带食蚜蝇与叶色草蛉主要以捕食蚜虫为主,因此时间生态位宽度要小于黑缘红瓢虫与异色瓢虫。
从时间生态位重叠值来看,桃蚜与桑白盾蚧和桃潜蛾的重叠值较大,分别为0.094 6和0.082 5,其次是桃蛀野螟与桃蛀果蛾,重叠值为0.069 8,继后是桑白盾蚧和桃潜蛾,重叠值为0.068 2,其余均较一致。重叠值越大则它们的时间占有量越相似,表明在时间维度上具有一定的同步性。黑缘红瓢虫与各主要害虫的时间生态位重叠值均较大,说明具有较强的同步性和跟随效应,对害虫有一定的抑制作用。异色瓢虫与桃蚜、桑白盾蚧的时间生态位重叠值也较大,可见异色瓢虫是这2种害虫的主要捕食者。
时间生态位相似性比例指数是测定物种在利用资源过程中的相似性指标,是对时间生态位重叠指数的进一步反映。从表 2可见,桃蚜与桑白盾蚧和桃潜蛾的时间生态位相似性比例指数值较大,分别为0.833 5和0.796 5,其次是桃蛀野螟与桃蛀果蛾,相似性比例指数值为0.731 3,继后是桑白盾蚧和桃潜蛾为0.719 4,其余均较小,说明这些害虫优势种在占据资源的时间生态位上有很大的相似程度,其余则不同。黑缘红瓢虫与害虫亚群落优势种的时间生态位相似性比例指数值较大,异色瓢虫与桃蚜、桑白蚧、梨小食心虫的相似性比例指数值也较大,说明这些天敌与各类害虫在时间上具有一定的同步性与跟随效应,这一结果进一步表明时间生态位重叠中所得的结论。
2.2.3 时、空二维生态位分析物种的存在与竞争离不开时间和空间,因此时、空二维生态位能够比较准确地反映共存与竞争的机制。由表 3可知,害虫优势种的时、空二维生态位宽度依次为:桑白盾蚧(0.578 7) >桃蚜(0.543 7) >桃潜蛾(0.516 0) >梨小食心虫(0.471 2) >桃蛀野螟(0.346 0)>桃蛀果蛾(0.315 7)。调查表明:桑白盾蚧不仅发生危害期要较早,而且几乎在桃树的各个部位均能发现它的寄生。桃蚜在桃树空间上的利用与发生危害期仅次于桑白盾蚧,可见它们对时间与空间资源的利用均较大。桃潜蛾主要以幼虫潜叶危害,而梨小食心虫主要加害桃树顶梢,因此这些害虫优势种的时、空二维生态位相对较小。天敌优势种的时、空二维生态位宽度依次为:黑缘红瓢虫(0.549 8)>异色瓢虫(0.476 9)>黑带食蚜蝇(0.440 6)>叶色草蛉(0.415 8)>小花蝽(0.353 1),由此可见,黑缘红瓢虫对害虫时间与空间资源的利用均较大,是桃园重要的天敌资源。
|
|
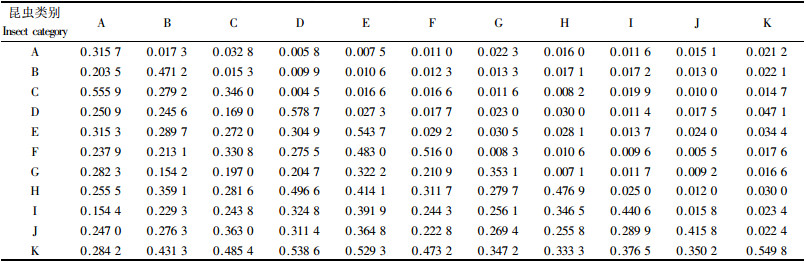
桃蛀野螟与桃蛀果蛾的时、空二维生态位重叠值最大(0.032 8),其次是桃蚜与桃潜蛾(0.029 2),桑白盾蚧和桃蚜的时、空二维生态位重叠值相对要小(0.027 3)。这显然是由于它们的生物学特性不同而致,桃蛀野螟与桃蛀果蛾均以幼虫蛀果危害,而桃蚜与桃潜蛾共同寄生于叶部进行刺吸或潜叶危害,它们在二维资源上具有类似的资源占有量,并且由于种群数量大而导致竞争激烈;桑白盾蚧主要寄生于2~3年生的枝条吸食危害,而桃蚜主要刺吸叶片,它们在资源的利用与竞争上是分离的,致使时、空二维生态位有一定的偏离。黑缘红瓢虫与各类害虫的时、空二维生态位重叠值皆较大,尤其与桑白盾蚧的重叠值最大(0.047 1),同样异色瓢虫与桑白盾蚧的重叠值也较大(0.030 0),由此可见,害虫与天敌的优势种群在时间上具有明显的同步性,在空间上有一定的同域性,对害虫有一定的调控能力。
从表 3可见,桃蛀野螟与桃蛀果蛾的时、空二维生态位相似性比例指数最大(0.555 9),其次是桃蚜与桃潜蛾(0.483 0),说明它们在时间资源的利用上和在空间资源的占据中有着较强的同步性和同域性。黑缘红瓢虫与各类害虫的时、空二维生态位相似性比例指数皆较大,异色瓢虫与桑白盾蚧、桃蚜的相似性比例指数也较大,说明它们在桃园生态环境中同时存在的机遇较大,共同占有相同空间的时间较长,有明显的跟随效应,这一结论进一步表明时、空二维生态位重叠中所得出的结论。
从表 1~3的结果不难看出,无论是时、空二维生态位宽度、重叠,还是相似性比例指数都小于时间、空间生态位宽度、重叠及相似性比例指数。可见环境变量之间不是绝对独立的,他们是相互依赖、相互作用和相互制约的,在一定的条件下共同影响着物种对资源的利用,使得物种对资源的利用程度比一维条件下相对要小,多维生态位的宽度、重叠,还是相似性比例指数也相应变小。
3 结论与讨论北京观光桃园昆虫群落的组成较为丰富,物种数为34种,隶属9目25科,其中植食性种类18种,天敌昆虫11种,中性昆虫5种。桃蛀果蛾、梨小食心虫、桃蛀野螟、桃潜蛾、桑白盾蚧、桃蚜、小花蝽、异色瓢虫、黑带食蚜蝇、叶色草蛉、黑缘红瓢虫等是桃园昆虫的优势种,桑白盾蚧和黑缘红瓢虫在桃园尤为突出。害虫亚群落在一定的区域和树上的布局等自然条件下,随着季节与物候的变迁,形成了自身发生、发展与衰退的自然规律,作为取食害虫的天敌亚群落,则随着害虫的增长形成了自身的消长规律。害虫亚群落有其栖境和空间生态位,而天敌亚群落也有一定的栖境和空间生态位,在桃园与它们的猎物和活动范围相互应。这就使得天敌亚群落在数量上必须与害虫亚群落发生空间分布相似,时间发生上同步,天敌亚群落才对害虫亚群落能够起到较好的调控作用。研究结果表明,瓢虫类天敌昆虫对桃园害虫亚群落具有较好的调控能力,黑带食蚜蝇、叶色草蛉、小花蝽也起到了一定的作用。蚧虫类、蚜虫类的危害尽管发生在整个桃树的生长过程中,但在时间与空间二维生态位上都受到黑缘红瓢虫与异色瓢虫的制约。在桃树整个生长过程中,昆虫群落优势度较高,表明害虫亚群落与天敌亚群落基本处于一定的平衡状态,但天牛类、潜叶类等一些生活习性较为隐蔽的害虫,常有点片发生危害的情况。因此,除了保护和利用桃园的自然天敌外,还应该辅以必要的人工释放天敌,或利用生物调节剂进行合理的调治,以保证桃园害虫亚群落与天敌亚群落的相对平衡与稳定,把桃园害虫优势种控制在经济允许受害水平之下(师光禄等, 1994)。
生态位是物种栖息、取食、繁殖等具有特定意义的场所,它反映的是物种对资源的利用情况。生态位定量描述的宽度和重叠,其实质是物种对各种资源的开发利用程度的定量测定,其结果不仅可以反映出群落中各物种对空间、时间、食物及其他环境资源的利用程度与关系,也为正确评判物种的属性提供了参考的依据。多维生态位的量化概念是指在时间条件下,物种对必须资源利用的多维空间,研究物种多维生态位随着时间的演变有助于了解物种的演替与进化,以及对其功能及地位的测定。时空生态位既是多维生态位的重要组分,又是多维生态位的特定反映形式(Bugg et al., 1994)。本文仅对桃园昆虫群落及其优势种的生态位进行了调查与分析,并结合优势种的生物学特性,明确了它们之间的相互关系,如果结合营养等其他环境因子,进行多维生态位的分析研究,将会较为深入地阐述天敌亚群落对害虫亚群落的调控机制,为桃园天敌的利用,以及害虫的生态调控提供更多的信息资源。
韩宝瑜, 张钟宁. 2000. 马尾松林直翅目昆虫群落的时空格局及其多样性和稳定性. 昆虫学报, 43(Sp.1): 143-150. |
黄保宏, 邹运鼎, 毕守东, 等. 2005. 梅园昆虫群落特征、动态及优势种生态位. 应用生态学报, 16(2): 307-312. DOI:10.3321/j.issn:1001-9332.2005.02.022 |
李昌根, 邹运鼎, 毕守东, 等. 2005. 管理水平对葡萄园节肢动物群落特征参数及目标害虫、天敌种群数量的影响. 应用生态学报, 16(12): 2365-2368. DOI:10.3321/j.issn:1001-9332.2005.12.028 |
师光禄, 刘素琪, 曹挥, 等. 2005. 枣园生态系统中害虫与天敌群落演替规律的研究. 应用生态学报, 16(11): 2130-2134. DOI:10.3321/j.issn:1001-9332.2005.11.024 |
师光禄, 刘素琪, 曹挥, 等. 2006a. 枣园害虫、捕食性和中性昆虫群落结构及动态的研究. 应用生态学报, 17(1): 80-86. |
师光禄, 王有年, 刘素琪, 等. 2006b. 枣麦间作枣园主要蚧虫复合种群结构及生态位研究. 应用生态学报, 17(7): 1234-1238. |
师光禄, 郑王义, 党泽普, 等. 1994. 果树害虫. 北京: 中国农业出版社.
|
万方浩, 陈常铭. 1986. 综防区和化防区稻田害虫-天敌群落组成及多样性的研究. 生态学报, 6(2): 159-170. |
袁忠林. 1999. 陇南桔园昆虫群落生态位及时序动态研究. 西北农业学报, 8(2): 49-52. |
赵志模, 郭依泉. 1990. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 12.
|
周洪旭, 郭建英, 万方浩. 2004. 转Cry1Ac+CpTI基因棉对棉田害虫及其天敌种群动态的影响. 昆虫学报, 47(4): 538-542. DOI:10.3321/j.issn:0454-6296.2004.04.022 |
邹运鼎, 毕守东, 周夏芝, 等. 2003. 桃园害虫及天敌群落动态研究. 应用生态学报, 14(5): 717-720. DOI:10.3321/j.issn:1001-9332.2003.05.015 |
Bugg R L, Waddington C. 1994. Using cover crops manage arthropod pests of orchards: a review. Agricultural Ecosystem and Environment, 50(1): 11-28. DOI:10.1016/0167-8809(94)90121-X |
Levins R. 1968. Evolution in changing environments. New Jersey: Princeton University Press, 120.
|