 2009, Vol. 45
2009, Vol. 45文章信息
- 郭小勤, 李犇, 阮晓赛, 娄永峰, 林新春, 汤定钦, 方伟.
- Guo Xiaoqin, Li Ben, Ruan Xiaosai, Lou Yongfeng, Lin Xinchun, Tang Dingqin, Fang Wei
- 利用ACGM分子标记研究10个毛竹不同栽培变种的遗传多样性
- Genetic Diversity of Ten Phyllostachys edulis Cultivars with ACGM Markers
- 林业科学, 2009, 45(4): 28-32.
- Scientia Silvae Sinicae, 2009, 45(4): 28-32.
-
文章历史
- 收稿日期:2008-05-06
-
作者相关文章



毛竹(Phyllostachys edulis)是浙江省也是全国最重要的经济竹种,在竹产业的发展中占据着极其重要的地位。由于毛竹的栽培面积极大,在长期的生产经营中产生了一系列稳定的变异类型,据记载有变型或栽培型12个,可见毛竹种内的形态变异多样性。但有关毛竹种内遗传多样性的研究,仅见Lai等(1997)对台湾毛竹林无性系遗传变异的研究,以及师丽华等(2002)用RAPD标记对毛竹种下等级7个变型的研究。
扩增的共有序列遗传标记(amplified consensus genetic marker,ACGM)是Brunel等(1999)根据一种植物中已知功能基因的保守编码序列设计引物对另一种植物进行PCR扩增且适合于比较作图的分子标记。利用该技术,Fourmann等(2002)开发出32个可以在拟南芥(Arabidopsis thaliana)和油菜(Brassica campestris)之间共用的ACGM标记,并利用这些标记构建了油菜的遗传连锁图谱,且比较了油菜种内4个亚种的遗传信息。Wang等(2005)利用已公布的水稻(Oryza sativa)2亚种(籼稻93-11和粳稻Nipponbare)的基因组序列,借助生物信息学的方法,开发了5 800多个候选的ACGM标记,并通过试验从中开发出173个在水稻中普遍适用的ACGM标记。以此为基础,结合小麦(Triticum aestivum)、大麦(Hordeum vulgare)和玉米(Zea mays)的cDNA/EST序列,卢泳全等(2006a)开发了38对禾本科通用型ACGM标记,试验表明,这些标记在小麦属和竹子中有一定的通用性(卢泳全等,2006b;董德臻等,2007),并可以有效地用于禾本科(Gramineae)几个物种间和不同属竹子间的遗传分类(Lu et al., 2006)。
利用与上述相似的方法,笔者开发了2对ACGM引物。本文运用这40对ACGM标记对毛竹的10个不同栽培变种及其2个近缘种进行分析,以期利用已知功能的分子标记探索毛竹的遗传多样性。
1 材料与方法 1.1 植物材料选取毛竹不同栽培变种及其2个近缘种进行试验(表 1)。供试材料都采自安吉竹种园。竹叶采集时间为2007年5月,所采集的样品均通过专家的形态学鉴定,采集的竹叶样品装入放有硅胶的采集袋,带回实验室,立即放入-70℃冰箱备用。
|
|

采用CTAB法提取植物叶片DNA。PCR反应体系总体积为20 μL,其中含50 ng模板DNA,上、下游引物各0.5 μmol·L-1,200 μmol·L-1 dNTP,1.5 mmol·L-1 MgCl2,1 U Taq DNA聚合酶,2 μL 10×PCR反应缓冲液。反应条件为:94 ℃预变性3 min;94℃变性45 s,55℃退火60 s,72 ℃延伸90 s,38个循环;72℃延伸10 min。对于扩增产物特异性差或没有扩增产物的引物进一步优化退火温度。部分引物组合至少重复2次以确保扩增结果的可靠性。扩增产物在3%的琼脂糖凝胶上电泳。回收目的条带,克隆后测序(上海美季生物技术有限公司)。
1.3 扩增结果的统计及遗传进化关系的分析以相同迁移位置记作一个扩增位点,记录每对引物在12个供试材料中PCR扩增获得位点的总数Nt;12个材料中扩增获得的位点数Nb,及多态性位点数Np。
在每个引物的相同迁移位点上,以"1"或"0"记录条带信息,其中"1"代表有扩增产物,"0"代表无扩增产物,建立一个数据矩阵。根据Nei's(1978)的公式计算遗传距离。把距离矩阵输入Popgene 32软件包中进行统计分析,再用TREEVIEW绘制树状图。
2 结果与分析 2.1 扩增结果及其序列分析利用40对ACGM引物对12个供试材料进行PCR扩增,结果共有5对引物在12个供试材料中均没有扩增产物(表 2),它们分别是GA7, GA8,GA22,GA25,GA34。35对引物可以在至少1个材料中得到PCR产物,其中27对(67.5%)引物在12个供试材料中表现出多态性。8对引物在供试材料间没有多态性位点,它们分别是GA1,GA6,GA16,GA20,GA21,GA23,GA32,GA38。对GA1引物分别从假毛竹、毛竹、佛肚毛竹、绿槽毛竹及厚壁毛竹中扩增出的条带进行测序,结果表明,来自假毛竹、毛竹、绿槽毛竹及厚壁毛竹间的序列完全相同,而来自佛肚毛竹的序列与其他4条序列仅在内含子的第57碱基处存在差别,即佛肚毛竹中是碱基T,其他均为碱基C,因此,GA1引物有望开发成SNP标记而将佛肚毛竹与以上4个栽培变种分开。
|
|
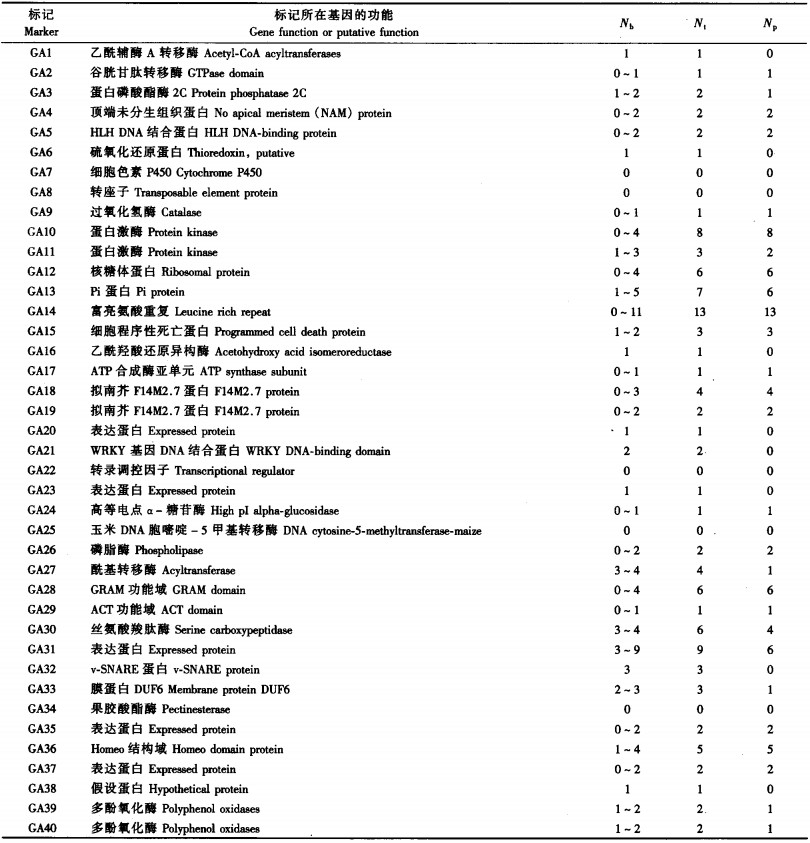
若按所用40对ACGM引物所在基因编码产物的功能进行分类,可将它们大致分为酶蛋白、转录调控因子及其他功能蛋白3类,其中以酶蛋白最多。从拷贝数来看,这些引物所在基因在水稻中均为单拷贝,但本文的结果显示,GA21,GA27, GA30,GA31, GA32, GA33共6对引物在所检测的12个样本中均能扩增出至少2条带(表 2),表现出多拷贝的特性。GA3,GA11, GA13,GA15, GA39,GA40共6对引物在某些供试样本中可扩增出单条带,而在另一些供试样本中扩增出多条带。如GA40引物在强竹、金丝毛竹、毛金竹、圣音毛竹、佛肚毛竹及龟甲竹中都能扩增出2条带,而在其他样本中均为单条带(图 1);进而对佛肚毛竹中所扩增出的条带进行测序,结果表明,这2条带确实来自与水稻同源的目标基因,且这2条带外显子部分联配得较好,而内含子则表现出很大的变异(图 2)。


|
图 2 GA40引物在佛肚毛竹中扩增产物序列的多重比对 Figure 2 Multiple alignments between the sequences of amplified products of primer GA40 in P. edulis cv. Ventricosa 横线表示相同的序列, 原点表示缺失的序列,方框内字母分别表示第1内含子的起始位置和第2内含子的终止位置。 Horizontal bars indicate the same sequence; the dots indicate lost sequence. GT and AG in pane indicate the starting position of the first intron and ending position of the second intron. |
利用40个ACGM标记在这些试验材料中扩增共得到89个多态位点。根据带型的不同,计算试验材料间的遗传距离。根据遗传距离,利用Popgene 32软件,得到10个毛竹不同栽培变种及其2个近缘种的聚类图(图 3)。从图 3中可知,遗传距离在0.4处时,可把12个试验材料分为3类,即毛金竹、假毛竹和不同栽培变种的毛竹。而在毛竹的2个近缘种中,假毛竹与毛竹各栽培变种的亲缘关系较近,毛金竹稍远。绿槽毛竹和黄槽毛竹的亲缘关系最近(遗传距离仅为0.060 1),这一结果与形态学分类结果一致。从聚类图中也可看出,花毛竹与强竹聚为一组,毛竹和厚壁毛竹聚为一组(遗传距离0.172 4)。其中圣音毛竹与毛竹其他栽培变种的亲缘关系较远。

|
图 3 10个毛竹不同栽培变种及2个近缘种的聚类结果 Figure 3 A dendrogram of ten cultivars and two affined species of P. edulis based on 40 ACGM markers |
以前在进行毛竹不同类型亲缘关系的研究中,所使用的分子标记都是随机标记。而ACGM分子标记是已知功能的基因标记,是从基因序列中开发的标记,排除了大量非编码序列的影响,它有助于开发带有已知功能基因的图谱,也有利于植物育种及分类学的研究,该标记更能体现功能基因在遗传本质上的差别。有研究表明,很多水稻的SSR标记并不能在其他植物中通用,而只能作为非特异的PCR标记,而水稻的ACGM标记可以在禾本科其他植物上通用(卢泳全等,2006b;董德臻等,2007)。与SSR标记相比,ACGM标记还具有一个特殊的优势,即它可直接反映基因间的变异。
本文的结果表明,在所利用的40对ACGM引物中,35对引物可以在至少1个材料中得到PCR产物(表 2)。其中27对(67.5%)引物在12个供试材料中表现出多态性。5对引物在供试材料中均没有扩增产物(表 2),从标记所在水稻基因的功能上看,这些标记的功能并不保守,如GA25所编码的玉米DNA胞嘧啶-5甲基转移酶,它可引起玉米DNA的甲基化,调节基因的功能,该蛋白家族除了在甲基转移酶结构域的同源性稍高外,其余序列的同源性都很低。但有些在禾本科其他物种中可以获得很好的扩增结果,如GA7,GA22,GA25在卢泳全等(2006b)所检测的12个小麦属材料和董德臻等(2007)所检测的13种竹子材料中都可获得特异条带,尽管GA7在所检测的样本中没有多态性位点。而GA8在董德臻等(2007)所检测的13种竹子材料中也未能获得扩增产物,在卢泳全等(2006b)所检测的12个小麦属材料中,有些种中最多可检测到4个扩增产物,说明竹子与小麦、水稻在该基因序列上存在差异。GA34在卢泳全等(2006)所检测的12个小麦属材料和董德臻等(2007)所检测的13种竹亚科材料中都未能获得扩增产物,说明该基因可能在禾本科各不同物种中存在特异性。
在能够获得PCR产物的35对引物中,有8对引物在供试材料间没有多态性位点(表 2),从标记所在基因的功能上看,这些标记多属于功能上十分重要的基因。例如,乙酰辅酶A是植物维持生命和生理功能的重要酶类,这些功能重要的基因要维持植物的正常生长,因此在毛竹不同栽培变种中的保守性很高。随机挑选以GA1引物分别从5个样本中扩增出的条带进行测序,结果表明,相同引物扩增出的序列间差异性很小。由此也证明了这2个基因在毛竹不同栽培变种及其近缘种中功能的保守性。
从凝胶电泳结果可得出,在毛竹的不同栽培变种中,最常见的多态性是缺失了扩增产物,即在一个或几个表型中无扩增产物,如GA29可在除强竹以外的11个供试材料中扩增出特异条带。这种现象可能是由于该表型中缺失该基因或者说该基因在引物位点发生了突变。另外还有一种多态性,即扩增产物大小间存在多态性。
该试验所用的12个材料都属于刚竹属,其中的10个材料为毛竹的不同栽培变种,而毛金竹和假毛竹是其近缘种。从外部形态看,近缘种及不同栽培变种的毛竹间存在明显的区别,仅用1对ACGM标记就可以将它们明显区别开来,有几对ACGM引物可在这2个近缘种中扩增出它们的特征谱带。但在毛竹的不同栽培变种中,大多数引物的扩增图谱都比较相似,说明从分子水平上也支持了将假毛竹和毛金竹作为单独的种,而毛竹各栽培变种间的亲缘关系较近,遗传背景较一致。从外部形态特征来看,黄槽毛竹和绿槽毛竹的区别主要在于秆的颜色,本文的研究结果也表明绿槽毛竹和黄槽毛竹的亲缘关系最近(遗传距离仅为0.060 1),这一结果与形态学分类结果相一致。同样,毛竹和厚壁毛竹的外部形态的区别主要是竹秆壁的厚度,其余形态特征趋于一致,本文的聚类结果中毛竹和厚壁毛竹被聚为一组(0.172 4)。在所测的10个毛竹不同栽培变种中,圣音毛竹与其他毛竹亲缘关系较远,这一结果与师丽华等(2002)的研究结果相吻合。
董德臻, 吴立成, 夏善勇, 等. 2007. ACGM标记在竹子中的通用性. 东北林业大学学报, 35(1): 4-6. DOI:10.3969/j.issn.1000-5382.2007.01.002 |
卢泳全, 汪旭升, 黄伟素, 等. 2006a. 基于水稻内含子长度多态性开发禾本科扩增共有序列遗传标记. 中国农业科学, 39(3): 433-439. |
卢泳全, 吴为人. 2006b. ACGM标记在小麦属中的通用性. 麦类作物学报, 26(5): 16-19. |
师丽华, 杨光耀, 林新春, 等. 2002. 毛竹种下等级的RAPD研究. 南京林业大学学报:自然科学版, 26(3): 65-68. |
Brunel D, Froger N, Pelletier G. 1999. Development of amplified consensus genetic marker (ACGM) in Brassica napus from Arabidopsis thaliana sequences of known biological function. Genome, 42: 387-402. DOI:10.1139/g98-141 |
Fourmann M, Barret P, Froger N, et al. 2002. From Arabidopsis thaliana to Brassica napus: development of amplified consensus genetic markers (ACGM) for construction of a gene map. Theoretical and Applied Genetics, 105: 1196-1206. DOI:10.1007/s00122-002-1040-z |
Lai C C, Hsiao J Y. 1997. Genetic variation of Phyllostachys edulis (Bambusoideae, Poaceae) in Taiwan based on DNA polymorphisms. Bot Bull Acad Sin, 38: 145-152. |
Lu Y Q, Ye Z H, Wu W R. 2006. Analysis of the phylogenetic relationships among several species of Gramineae using ACGM markers. Acta Genetica Sinica, 33(12): 1127-1131. DOI:10.1016/S0379-4172(06)60151-0 |
Wang X S, Zhao X Q, Zhu J, et al. 2005. Genome-wide investigation of intron length polymorphisms and their potential as molecular markers in rice (Oryza sativa L.). DNA Research, 12(6): 417-427. |