 2009, Vol. 45
2009, Vol. 45文章信息
- 卢立华, 蔡道雄, 贾宏炎, 何日明.
- Lu Lihua, Cai Daoxiong, Jia Hongyan, He Riming
- 南亚热带7种林分凋落叶养分含量的年动态变化
- Annual Variations of Nutrient Concentration of the Foliage Litters From Seven Stands in the Southern Subtropical Area
- 林业科学, 2009, 45(4): 1-6.
- Scientia Silvae Sinicae, 2009, 45(4): 1-6.
-
文章历史
- 收稿日期:2008-08-28
-
作者相关文章


凋落物在森林生态系统养分循环中担负着连接森林与土壤的“纽带”作用,它对于维持土壤地力,保持森林生态系统内物质的生物循环和养分平衡起到了重要作用(张万儒等,1986)。因此,有关凋落物养分研究的报道甚多(姚瑞玲等,2006;林瑞余等,2002;翁轰等,1993;田大伦等,1994;沈善敏等,1993)。
森林凋落物以凋落叶为主,方晰等(2005)对杉木(Cunninghamia lanceolata)凋落物的研究结果表明凋落叶占凋落物总量的69.8%;Arunachalam等(1998)对印度亚热带湿润次生林凋落物的研究结果表明凋落叶占凋落物总量的78%~88%;邹碧等(2006)对马占相思(Acacia mangium)、大叶相思(Acacia auriculaeformis)、湿地松(Pinus elliottii)及荷木(Schima superb)的研究结果表明,凋落叶占凋落物总量的83.2%~93.7%;卢立华等(2008)对米老排(Mytilaria laosensis)等6种人工林凋落物的研究结果表明,凋落叶占凋落物总量的61.32%~78.49%。可见,凋落叶在森林生态系统的养分循环中扮演了主要角色。此外,陆瑾(2006)对广玉兰(Magnolia grandiflora)的研究结果表明,凋落叶的养分浓度变化与活叶一致, 活叶浓度高, 对应凋落叶的浓度也高。因此,研究凋落叶养分的动态变化不仅能反映凋落物的养分动态变化规律及养分循环情况,也能了解林分的养分动态变化状况。本研究对杉木、马尾松(Pinus massoniana)、米老排、红椎(Castanopsis hystrix)、火力楠(Michelia macclurei)和西南桦(Betula alnoides) 6种南亚热带人工林及天然次生林凋落叶的养分进行动态变化分析,在该6种人工林中,仅有学者对马尾松和杉木的凋落物养分做过一些研究(杨细明,2002;田大伦等,1994),其余4种阔叶人工林凋落物的养分研究尚属空白。开展该研究可了解和掌握这些林分的养分动态变化状况及规律,从而更好地对林分进行科学的经营与管理,维持和提高地力,为实现人工林的可持续经营提供依据。
1 研究区概况研究在广西壮族自治区凭祥市中国林业科学研究院热带林业实验中心伏波实验场(21°57′47″—22°19′27″N,106°39′50″—106°59′30″E)进行,该区属南亚热带季风型半湿润—湿润气候区,太阳总辐射439.6 kJ·cm-2a-1, 年日照时数1 218~1 620 h, 年平均气温20.5~21.7℃,≥10℃年活动积温6 000~7 600℃; 年均降水量1 200~1 500 mm。干湿季节明显,4—9月为雨季,10月至翌年3月为旱季,雨热同季的气候特点对植物生长十分有利。研究区海拔650 m,属低山丘陵地貌,土壤为花岗岩发育的山地红壤,土层深厚、疏松,质地为轻壤土或重壤土,坡度为25°。主要灌木有山苍子(Litsea cubeba)、野牡丹(Melastoma candidum)、桃金娘(Rhodomyrtus tomentosa)大沙叶(Aporosa chinensis)和鹅掌柴(Schefflera octophylla);主要草本有五节芒(Miscanthus floridulus)、铁芒箕(Dicranopteris linearis)、蔓生莠竹(Microstegium ciliatum)、乌毛蕨(Blechnum orientale)和野香茅(Cymbopogon tortilis)。
2 研究方法在研究区东南坡海拔550~600 m处的杉木、马尾松、米老排、红椎、火力楠和西南桦人工纯林及天然次生林中各选择3块20 m×30 m的样地,样地林木生长良好,郁闭度为0.85~0.9。在每个样地中随机布设5个1 m×1 m的塑料纱网收集框, 用木桩固定于距地面30 cm高处,每月的15号定期收集凋落物1次。将每种林分3块样地的凋落物混匀,然后,对收集到的凋落物捡除非目的树种凋落物,按叶、枝、杂分类并分别称重。每月采用四分法选取凋落叶300 g作为分析样。将分析样烘干后粉碎装瓶,用于测定N,P,K,Ca及Mg含量。测定方法:N用半微量凯氏法测定,P用钼蓝比色法测定,K用火焰光度计法测定,Ca和Mg用原子吸收分光光度计法测定。凋落物收集时间为2006-05—2007-04,试验期间的气候因子见表 1。
|
|
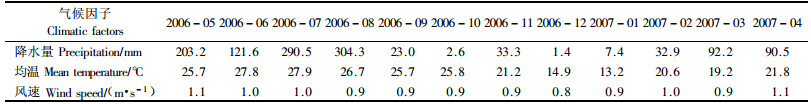
从表 2可得到7种林分凋落叶N含量的年变幅。在林木生长高峰期4—9月,凋落叶N含量相对较高,在林木缓长或休眠期的1,2,3,10,11与12月,凋落叶N含量相对较低。说明凋落叶N含量与林木的生长成正相关。4—9月林木的新陈代谢较为旺盛,林木为了满足其生长的需要,必须从土壤中吸收及从老器官中转移更多的N以满足其生长之需,而叶作为光合作用的主要器官,是物质与能量交换的重要场所,也是植物养分变化最为敏感的器官,植株养分的变化首先表现在叶片上;另一方面这段时间叶片凋落的周期缩短,加之这一时段为雨季,风雨较大,易导致部分叶片在没有完成N的正常迁移前掉落,这些因素导致了林分凋落叶N含量较高。
|
|
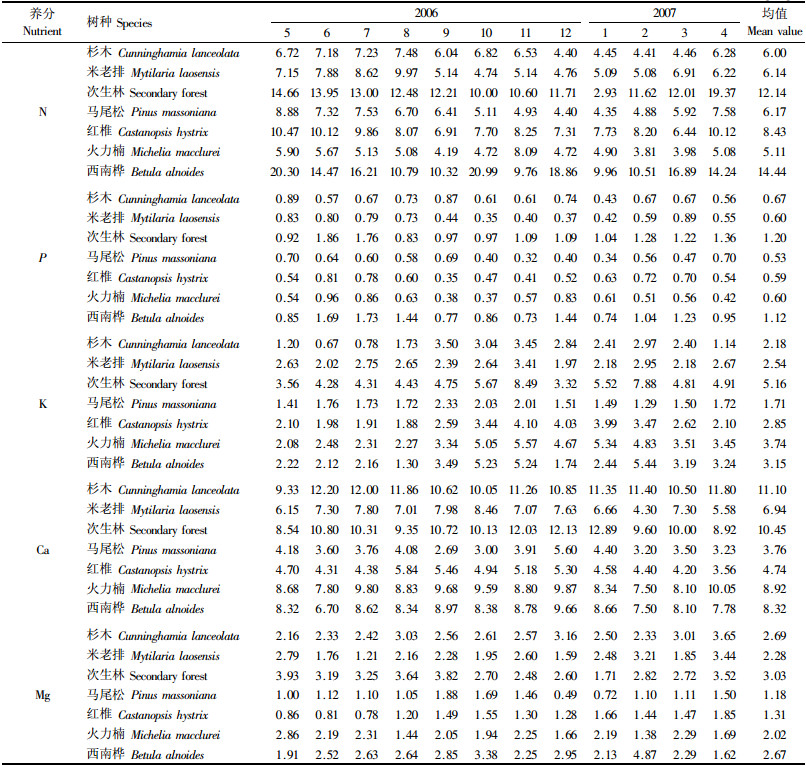
从表 2可见,N年平均含量表现为西南桦>次生林>红椎>马尾松>米老排>杉木>火力楠,以西南桦最高,达14.44 g·kg-1,火力楠最低,仅5.11 g·kg-1,高低相差1.83倍,马尾松、米老排和杉木的凋落叶年平均N含量较接近,分别为6.17,6.14和6.00 g·kg-1。
研究发现,常绿树种米老排、红椎、火力楠、杉木、马尾松凋落叶的N含量比落叶树种低。次生林的树种构成复杂,其林分凋落叶是多树种凋落叶的混合体,虽以常绿树种为主,但也不乏落叶树种,故其林分凋落叶N含量比落叶纯林低,而比常绿纯林高。产生差异的原因除不同树种构成叶片的组分含量不同外,还与树种的生理生态过程及N的转移再利用能力等有关,降雨及露珠的淋洗对凋落叶N含量也会产生一定的影响。
3.2 凋落叶P含量动态变化从表 2可见,不同树种凋落叶在不同月份的P含量以针叶林和常绿阔叶林全年波动较小,落叶林及次生林波动较大。
米老排P含量以3月最高,达0.89 g·kg-1,5—8月亦维持在较高水平,为0.73 ~0.83 g·kg-1,其他月份相差不大,且都维持在较低的水平,为0.35~0.59 g·kg-1;马尾松4—9月凋落叶P含量较高,为0.58~0.70 g·kg-1,1,2,3,10,11与12月都较低;杉木以5和9月较高,分别达0.89和0.87 g·kg-1,1月最低,仅0.43 g·kg-1,其他月份较为接近,为0.56~0.74 g·kg-1。本研究中,红椎、火力楠、西南桦和马尾松均为热带树种,对水热特别敏感,高温多雨对其生长及代谢有利,而林木要维持高的生长量,必须有高的P素营养作为保证,故其凋落叶的P含量也较高;米老排虽也属热带树种,但其相对喜荫,因此,其速生期要比红椎、火力楠和西南桦提前,3月份即进入速生期;而杉木为亚热带树种,俞新妥等(1982)的研究结果表明,气温过高,杉木的蒸腾作用大,树木体内水分亏缺严重,细胞分裂延长,林木的生长减弱,但温度过低同样影响其生理活动,对杉木的生长也不利,本研究中杉木凋落叶P含量的变化与其生长变化趋势较一致,故杉木凋落叶P含量的最高月出现在5和9月(杉木生长较快的月份)。综上可见,7种林分凋落叶P含量的动态变化基本上与林分的生长动态变化趋势一致,即凋落叶P含量与林木生长成正相关。
P年平均含量表现为次生林>西南桦>杉木>米老排>火力楠>红椎>马尾松,以次生林最高,达1.20 g·kg-1,马尾松最低,仅0.53 g·kg-1,高低相差1.26倍,西南桦的P含量较高,达1.12 g·kg-1,其余4个树种都较低,且很接近,为0.59 ~0.67 g·kg-1。
3.3 凋落叶K含量动态变化从表 2可见,杉木凋落叶K含量4—8月较低,为0.67~1.73 g·kg-1,其他月份相对较高,为2.40~3.50 g·kg-1;红椎、火力楠的动态变化趋势较相似,都是3—9月较低,分别为1.88~2.62和2.08~3.51 g·kg-1,1,2,3,10,11及12月较高,分别为3.44~4.10和4.67~5.57 g·kg-1;西南桦在1,5,6,7,8及12月都较低,为1.30~2.44 g·kg-1,2—4及9—11月都较高,为3.19~5.44 g·kg-1;天然次生林在3—9和12月较低,为3.32~4.91 g·kg-1,在1,2,10和11月较高,为5.52~8.49 g·kg-1;而马尾松、米老排凋落叶的K含量没有表现出明显的高低时间段,马尾松的年变幅为1.29~2.33 g·kg-1,米老排为1.97 ~3.41 g·kg-1。
7种林分凋落叶K含量总体上表现为1—4及9—12月较高,4—8月较低,且这几个月中凋落叶K含量的差异不大。说明在林木生长高峰期多数树种凋落叶的K含量都较低,而在林木缓长期或休眠期,凋落叶K含量反而较高。林木中K较活跃,很容易从老叶向嫩叶或其他幼嫩的器官迁移进行再利用,由于林木在速生期对K的需求量会增加,林木为了满足生长的需要,除从土壤中吸收更多的K元素外,还会从老营养器官中尽可能多地向嫩营养器官中迁移再利用K元素。而在秋冬季,林木生长缓慢,甚至进入了休眠状态,生理活动较弱,对K的需求量也相对减少,养分的内迁移作用随之减弱,导致了凋落叶K含量在林木生长旺盛期较低,而缓长或休眠期反而高。红椎与火力楠这2个树种的生长特点较相似,它们凋落叶的含K量动态变化趋势亦较为一致。秦国峰(2000)对马尾松的研究表明:马尾松在我国南亚热带地区几乎全年无明显高、粗生长休眠期,故它的凋落叶含K量也没有明显的高低时间段之分,综上可见,凋落叶的K含量与林木生长呈负相关。
K年平均含量表现为次生林>火力楠>西南桦>红椎>米老排>杉木>马尾松,以天然次生林最高,达5.16 g·kg-1,马尾松最低,仅1.71 g·kg-1,高低相差达2.02倍。
3.4 凋落叶Ca含量动态变化从表 2可见,7种林分凋落叶Ca含量年变幅不大。植物中的Ca大部分存在于叶内,而大量的Ca以果胶钙形式被永久地固定于细胞壁的中胶层,故Ca在植物体内不容易移动和重新分配。由于不同月份中同一树种构成叶片的组分及含量相对稳定,尽管在不同月份中,因受气候及生长差异的影响,对Ca的吸收量会有所不同,导致凋落叶Ca含量在各个月中会有些波动,但波动不会太大,故同一树种的凋落叶在不同月份中Ca含量变幅最小。但是,不同林分凋落叶的Ca含量差异却较明显。7种林分凋落叶Ca年平均含量表现为杉木>天然次生林>火力楠>西南桦>米老排>红椎>马尾松,以杉木最高达11.1 g·kg-1,马尾松最低仅1.71 g·kg-1。这主要是由树种遗传特性决定的,如马尾松为厌Ca喜酸树种,它在生理过程中需Ca量相对较低,故它在生长过程中对Ca的吸收也最少,导致其凋落叶中Ca含量最低。
3.5 凋落叶Mg含量动态变化从表 2可见,7种林分凋落叶Mg含量随月份的波动较大,基本上为波浪式。
杉木以4月最高,达3.65 g·kg-1,5月最低,为2.16 g·kg-1,最高月比最低月高了68.89%;其余月份以1,2,6,7,9与10月份的Mg含量较低,相差也较小,为2.33~2.61 g·kg-1;3,8与12月较高,分别为3.01,3.03与3.16 g·kg-1,相差也不大。
米老排以4月份含量最高,为3.44 g·kg-1,7月最低,仅1.21 g·kg-1,高低相差达1.84倍,3,6,8,9,10和11月含量都不高,为1.59~2.28 g·kg-1。
次生林以5月份最高,达3.93 g·kg-1,1月最低,仅1.71 g·kg-1,高低相差1.3倍,4—9月处于较高水平,含量为3.19~3.93 g·kg-1,2,3,10,11及12月较低,含量为2.48~2.82 g·kg-1。
马尾松以9月份最高,为1.88 g·kg-1,12月最低,仅0.49 g·kg-1,高低相差达2.84倍,10与11月处于较高水平,分别为1.69与1.46 g·kg-1,1月份很低,为0.72 g·kg-1,2,3,4,5,6,7和8月都稳定在1.00~1.11 g·kg-1之间。马尾松凋落叶的Mg含量为7种林分中之最低,也是月份间相差倍数最大的1个树种,这与其喜酸厌碱的特性有关。
红椎以4月最高,为1.85 g·kg-1,7月最低,为0.78 g·kg-1,5和6月较低,分别为0.86和0.81 g·kg-1,其他月较高,相差也较小,为1.20~1.66 g·kg-1。
火力楠以5月最高,为2.86 g·kg-1,2月最低,为1.38 g·kg-1,高低相差1倍多,4,8,10和12月都较低,且相差不大,为1.44~1.94 g·kg-1;其余月份较高,相差也较小,为2.05~2.31 g·kg-1。
西南桦以2月最高,为4.87 g·kg-1,4月最低为1.62 g·kg-1,高低相差达2倍,10月份较高,达3.38 g·kg-1,5月较低为1.91 g·kg-1,1,3,6,7,8,9,11与12月含量为2.13~2.85 g·kg-1。
Mg年平均含量表现为次生林>杉木>西南桦>米老排>火力楠>红椎>马尾松,以次生林的年平均含量为最高,达3.03 g·kg-1,马尾松最低,仅1.18 g·kg-1,高低相差1.57倍。
4 结论与讨论对广西凭祥市中国林业科学研究院热带林业实验林中心7种林分凋落叶养分动态变化研究结果表明,在热带南亚热带地区,林生生长与林分凋落叶的N和P含量成正相关,与K含量成负相关,首次明确提出了凋落叶养分含量与林木生长具有明显的相关性结论。这一研究结论与林成芳等(2005)对福建柏人工林凋落物养分动态特征研究中不同月份的凋落叶N,P和K含量结果基本一致,同时,也与李发林等(2002)对龙眼果树凋落叶N,P和K含量季节变化动态结果相吻合。但与薛立等(2001)对名古屋风景林凋落物和凋落叶养分含量季节动态的研究结论有相左之处,是否因其研究区域与本研究区域的气候带差别明显所致,还有待继续研究。
森林凋落物分解过程中每年释放的营养元素可满足69%~87%的森林生长需求量(Warning et al., 1985)。森林凋落物以凋落叶为主,了解林分凋落叶养分含量的动态变化规律,对于深入了解森林的养分循环及利用机制具有重要意义,同时,可为人工林树种的合理布局和科学经营与管理提供依据。如西南桦的凋落叶N和P含量都较高,说明它对土壤N和P的自肥能力也较强,反过来也说明它对N和P的需求量也较多,故在造林时应选择N和P含量较丰富的土壤,在对其进行施肥时也应以N和P肥为主。马尾松的凋落叶P,K,Ca及Mg含量都为最低,N的含量也较低,这与其喜酸厌碱的特点相吻合,同时也说明它对土壤的自肥能力最差,也是本研究树种中最耐瘠薄的一个树种,故在条件不是特别差的立地上经营其人工林一般可不必施肥。另外,马尾松为强阳性树种,故在造林树种布局上可将它安排在土壤肥力相对较低、强酸性、阳光充足的山坡上部、山脊等地段种植。红椎的凋落叶N,P,K,Ca及Mg的含量也都较低,与其他树种比较也相对的耐瘠薄,且其幼林期较耐荫,因此,可安排在阴坡,土壤相对瘠薄的林地中造林。其他几个树种对肥力的要求差不多,但米老排较耐潮湿,可安排在沟壑边或低洼地种植。此外,通过对林分凋落叶养分进行更深入的研究分析,应能提出一种更便利、准确的林木营养诊断方法,这还有待在今后的研究中解决。
森林凋落物的养分必须通过凋落物的分解才能回归土壤进行再利用,而凋落物分解速率的高低在很大程度上决定了一个生态系统(尤其是森林生态系统)生产力的高低(Adriend et al., 2001)。因此,开展凋落物分解速率、分解过程中的养分动态与凋落物质量、生境、生物以及气候之间的关系,尤其是土壤生物(微生物和动物等)对凋落物分解影响等的研究十分必要和迫切。此外,凋落物分解与一些环境热点问题的关系(如多样性丧失对凋落物分解过程的影响以及凋落物分解在碳循环和氮循环中的作用等)也是今后需要研究的领域。
方晰, 田大伦, 项文化, 等. 2005. 杉木人工林凋落物量及其分解过程中碳的释放率. 中南林学院学报, 25(6): 12-16. DOI:10.3969/j.issn.1673-923X.2005.06.002 |
李发林, 黄炎和, 蔡志发, 等. 2002. 龙眼果树凋落物和凋落叶N、P、K含量季节变化动态. 福建农业学报, 17(2): 108-111. DOI:10.3969/j.issn.1008-0384.2002.02.011 |
林成芳, 李震, 牛志鹏, 等. 2005. 福建柏人工林凋落物的养分动态特征. 福建农林大学学报:自然科学版, 34(1): 63-66. |
林瑞余, 陈银秀, 黄荣臻, 等. 2002. 杉木观光木混交林凋落物养分特征及动态变化. 东北林业大学学报, 30(1): 17-23. DOI:10.3969/j.issn.1000-5382.2002.01.005 |
卢立华, 贾宏炎, 何日明, 等. 2008. 南亚热带6种人工林凋落物的初步研究. 林业科学研究, 21(3): 346-352. DOI:10.3321/j.issn:1001-1498.2008.03.012 |
陆瑾. 2006. 广玉兰凋落叶NPK元素的营养转移. 安徽农业科学, 34(16): 3897-3898. DOI:10.3969/j.issn.0517-6611.2006.16.012 |
秦国峰. 2000. 马尾松改良及培育. 杭州: 浙江大学出版社, 2-5.
|
沈善敏, 宇万太, 张璐, 等. 1993. 杨树主要营养元素内循环及外循环研究Ⅰ——落叶前后养分在植株体内外的迁移和循环. 应用生态学报, 4(1): 27-31. |
田大伦, 谌小勇, 康文星, 等. 1994. 杉木林生态系统凋落物分解量及养分释放规律. 哈尔滨: 东北林业大学出版社, 146-153.
|
翁轰, 李志安, 屠梦照, 等. 1993. 鼎湖山森林凋落物量及营养元素含量研究. 植物生态学与地植物学学报, 17(4): 299-304. DOI:10.3321/j.issn:1005-264X.1993.04.012 |
薛立, 薛达, 罗山. 2001. 名古屋风景林凋落物和凋落叶养分含量季节动态的研究. 植物生态学报, 25(3): 359-365. DOI:10.3321/j.issn:1005-264X.2001.03.016 |
杨细明. 2002. 马尾松人工林凋落物分解及养分释放规律. 福建林学院学报, 22(1): 1-3. DOI:10.3969/j.issn.1001-389X.2002.01.001 |
姚瑞玲, 丁贵杰, 王胤. 2006. 不同密度马尾松人工林凋落物及养分归还量的年变化特征. 南京林业大学学报:自然科学版, 30(5): 83-86. |
俞新妥, 陈存及, 郭先魁. 1982. 杉木不同种源树高生长昼夜节律的研究. 福建林学院学报, (2): 25-27. |
张万儒, 许本彤. 1986. 森林土壤定位研究方法. 北京: 中国林业出版社.
|
邹碧, 李志安, 丁永祯, 等. 2006. 南亚热带4种人工林凋落物动态特征. 生态学报, 26(3): 715-721. DOI:10.3321/j.issn:1000-0933.2006.03.011 |
Adriend C F, Andrew S A, Evan H D, et al. 2001. Forest litter production, chemistry, and decomposition following two years of free air CO2 enrichment. Ecology, 82(2): 470-484. |
Arunachalam A, Arunachalam K M, Pandey H N, et al. 1998. Fine litterfall and nutrient dynamics during forest regrowth in the humid subtropics of north-eastern India. Forestry Ecology and Management, 110: 209-219. DOI:10.1016/S0378-1127(98)00282-5 |
Warning R H, Schlesinger W H. 1985. Forest ecosystems: concepts and management. New York: Academic Press, 181-210.
|