 2009, Vol. 45
2009, Vol. 45文章信息
- 金松恒, 徐礼根, 李雪芹, 王俊刚, 朱澜, 贾晓琳.
- Jin Songheng, Xu Ligen, Li Xueqin, Wang Jungang, Zhu Lan, Jia Xiaolin
- 长期高温胁迫对高羊茅光合特性和抗氧化酶活性的影响
- Effects of Long-term High Temperature Stress on Photosynthetic Characteristics and Antioxidant Activity in Festuca arundinacea
- 林业科学, 2009, 45(3): 155-159.
- Scientia Silvae Sinicae, 2009, 45(3): 155-159.
-
文章历史
- 收稿日期:2007-12-13
-
作者相关文章


2. 浙江大学生命科学学院 杭州 310058;
3. 浙江林学院天目学院 临安 311300
2. College of Life Sciences, Zhejiang University Hangzhou 310058;
3. Tianmu College, Zhejiang Forestry University Lin'an 311300
高温已成为影响植物正常生长的主要因素之一,尤其是我国南方的夏季高温已经成为制约许多植物生长和发育的主要环境因子,也给林草生态安全和植被景观带来极大隐患。高羊茅(Festuca arundinacea),是常见的冷季型草坪草中绿期最长、应用最广的草种之一(周嘉友等,1998;韩春梅等,2006),在气候冷凉地区能周年常绿,在长江中下游气候过渡区绿期也相对较长,但在高温季节常出现大面积枯黄现象。在长江以南地区,因长期处于夏季高温的影响,普遍存在难以越夏的问题。因此高温胁迫成为高羊茅在我国南方草坪发展的主要限制因子。目前, 国内外已有许多关于高羊茅耐热性方面的研究(何亚丽等, 2002;徐胜等,2008;Cui et al., 2006;Jiang et al., 2001;Zhao et al., 2008)。高温胁迫能明显降低高羊茅的光合速率和PSII活性(Cui et al., 2006),而通过叶面喷施N肥可以明显减缓高温下高羊茅叶片净光合速率、气孔导度、水分利用效率和PSII最大光化学效率的下降(Zhao et al., 2008)。通过热锻炼能提高高羊茅在忍受高温过程中的耐热性(徐胜等,2008),对高羊茅叶片喷施水杨酸或Ca2+能够提高叶片超氧化物歧化酶和过氧化氢酶的活性从而提高耐热性(何亚丽等, 2002;Jiang et al., 2001)。前人对高羊茅等草坪草耐热性的研究大多集中在短期高温对叶片光合速率、抗氧化酶活性、膜脂过氧化等方面,对于长期处在高温环境过程中草坪草的光合作用、叶绿素荧光特性和光能分配等研究未见报道。本试验根据南方夏季高温对草坪草的实际情况,研究长期高温胁迫对高羊茅气体交换参数、叶绿素荧光参数、能量分配、叶绿素含量和抗氧化酶等的影响。以期了解长期高温对高羊茅的作用机制。
1 材料与方法 1.1 供试材料与培养高羊茅品种为猎狗5号(Festuca arundinacea cv. ‘Houndog 5’)。种在不同的花盆中,置光照培养箱内培养。光强为600 μmol·m-2 s-1, 温度为20 ℃/18 ℃(昼/夜),光周期为12 h,相对湿度为70%~75%。待植株长到5叶1心时,将其中一半高羊茅转移至另一光照培养箱进行高温处理,处理的最终温度为35 ℃/26 ℃(昼/夜),处理时昼温度从20 ℃开始每小时温度升高2 ℃,夜温由培养箱自行调节,其余条件不变,原培养箱的一半作为对照。处理第30天测定叶片气体交换参数和叶绿素荧光参数,每次均测定第2完全展开叶片(自上而下),测定后取相同的叶片用于测定叶绿素含量和抗氧化酶类等指标。
1.2 气体交换和叶绿素荧光参数测定用便携式光合测定系统(LiCor-6400,USA)和荧光叶室(6400-09)同时测定气体交换和叶绿素荧光参数,每次测定至少重复4次以上;所有测定时的气体流量均为500 μmol·s-1;测定条件为CO2浓度为(385±5) μmol·mol-1;Fo和Fm的是在叶片暗适应15 min后进行,用测量光先测定Fo,再用饱和的红蓝光照射,获取最大荧光(Fm),Fv/Fm=(Fm-Fo)/Fm;光响应曲线测定时光强设置为2 000,1 600,1 200,1 000,800,500,200,100,50,20,0 μmol·m-2s-1。其他的叶绿素荧光参数计算公式按Genty等(1989)的公式计算:PSⅡ的实际量子产额(ΦPSⅡ)=(F′m-Fs)/F′m,开放PSII反应中心激发能捕获效率F′v/F′m= (F′m-F′o)/F′m;光化学猝灭系数qP= (F′m-Fs)/(F′m-F′o);非光化学猝灭NPQ = Fm/F′m-1;电子传递速率(ETR)按Bilger和Bjôrkman的方法测定(1990)。PSⅡ吸收的光能分成3部分,在光强为800 μmol·m-2s-1时,用于PSII光化学反应(P)、天线耗散(D)和过剩光能(Ex)分别所占的比率根据Demmig-Adams等(1996)的公式计算。
1.3 CO2响应曲线测定利用CO2钢瓶完成,测定时的气体流量均为500 μmol·s-1,光照强度为1 200 μmol·m-2s-1。参比室CO2浓度设置按400,300,200,100,50,400,400,600,800,1 000,1 200,1 600 μmol·mol-1,每浓度适应4~6 min。测定由LI-6400内置程序自动完成。Rubisco最大羧化效率(Vc, max)和RuBP最大再生速率(Jmax)用Photosyn Asisstant软件计算。
1.4 NPQ的诱导和恢复动力学测定按照Quick等(1989)的方法进行。每分钟读取1个数值,由仪器自动完成。待仪器读取第15个值时关闭作用光。高能态猝灭(qE)和光抑制猝灭(qI)按照Thiele等(1997)的方法计算。
1.5 叶绿素和可溶性蛋白含量的测定参照Huang的方法(2004)。可溶性蛋白质的含量按考马斯亮蓝染色法测定,以牛血清白蛋白作标准曲线,计算可溶性蛋白含量。
1.6 抗氧化酶类活性的测定超氧化物歧化酶(SOD)活性根据Giannopolitis等(1977)的方法以抑制NBT光化学还原的能力来测定。过氧化物酶(POD)活性测定采用Braber(1980)的方法,根据470 nm波长下吸光度的增加来计算。过氧化氢酶(CAT)的活性根据Jiang等(2001)的方法,通过测定H2O2在240 nm下的降解速度来计算。抗坏血酸过氧化物酶(APX)活性根据Nakano等(1981)的方法测定。谷胱甘肽还原酶(GR)活性采用Foyer等(1976)的方法测定。
1.7 丙二醛(MDA)含量的测定含量测定参照Huang等(2004)的方法,用硫代巴比妥酸(TBA)比色法测定MDA的形成。
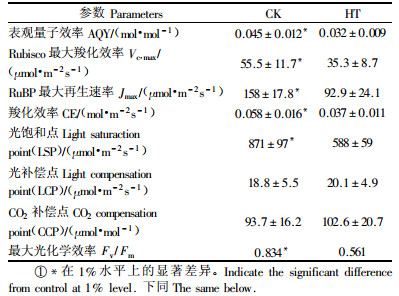
2 结果与分析 2.1 长期高温对高羊茅气体交换参数的影响在各个光强和CO2浓度下,高温胁迫均使高羊茅的光合速率下降,在高光强和高CO2浓度下显著(图 1)。利用Photosyn Asisstant软件对Pn-PFD和Pn-Ci响应曲线分析表明,长期高温处理使高羊茅表观量子效率AQY,Rubisco最大羧化效率Vc, max、RuBP最大再生速率Jmax、羧化效率CE和光饱和点分别下降29.9%,37.4%,41.2%,36.2%和38.9%。而光补偿点和CO2补偿点则分别上升6.9%和9.5%.

|
图 1 高温胁迫对高羊茅净光合速率(Pn)光强响应曲线和CO2响应曲线的影响 Figure 1 The effects of long-term high temperature stress on the response curves of net photosynthetic rate to light intensities and CO2 concentrations in tall fescue leaves |
在长期高温胁迫下,高羊茅的最大光化学效率Fv/Fm明显降低(表 1)。在任何光强下受高温胁迫的高羊茅的PSII反应中心激发能捕获效率(F′v/F′m)都更低(图 2A)。光化学淬灭(qP)随着光强呈直线下降(图 2B)。高温胁迫下,qP的下降幅度比对照的更大。qP值的大小反映的是原初电子受体QA的氧化还原状态和PSⅡ开放中心的数目(Maxwell et al., 2000),高温胁迫下qP值越低表明说明质体醌(PQ)还原程度随高温胁迫而增加。PSII实际量子效率ФPSⅡ反映PSⅡ反应中心在环境胁迫中有部分关闭情况下的实际原初光能捕获效率, 也是实际的PSⅡ反应中心进行光化学反应的效率,其值越小, 说明电子传递活性越小(Maxwell et al., 2000)。在每个光强下,高温胁迫下的ΦPSⅡ都比对照低得多(图 2C)。根据ΦPSⅡ计算线性电子传递速率(ETR)可知:高温胁迫下的ETR比对照的小得多,饱和ETR值大约只有对照的55%左右。而且高温处理下,高羊茅的ETR饱和光强只有500 μmol·m-2s-1左右,而对照则为1 000 μmol·m-2s-1左右(图 2D)。
|
|

|
图 2 高温对高羊茅叶绿素荧光参数和电子传递速率的影响 Figure 2 Effects of high temperature on chlorophyll fluorescence parameters and electron transport rate in tall fescue leaves |
NPQ的光强响应曲线,结果如图 3A所示。在低光强下,高温处理后高羊茅的NPQ和对照的无明显差异,但光强超过500 μmol·m-2s-1 时,高温处理下高羊茅的NPQ随光强上升缓慢,而对照则仍然迅速上升,饱和NPQ值显著高于高温处理的高羊茅。根据荧光暗恢复动力学并分别计算出NPQ的组分高能态猝灭(qE)和光抑制猝灭(qI)。qE是NPQ的主要组分,正常条件下占80%以上。qE的随光强变化的趋势基本和NPQ一致(图 3B),说明高温处理下NPQ的下降主要是由于qE的下降引起的。在光强高于200 μmol·m-2s-1时,高温处理的高羊茅的qI比对照高得多(图 3C)。qI的升高表明高温处理增加高羊茅对高光强的敏感性。

|
图 3 长期高温对NPQ (A)及其主要组分qE (B)和qI (C)的影响 Figure 3 Effects of long-term high temperature on the total NPQ and its distinct components:qE and qI in the leaves of tall fescue plants |
将叶片吸收的光能分为3部分(Demmig-Adams et al., 1996):一是用于PSⅡ光化学反应的部分P, 二是通过天线热耗散的部分D,三是PSⅡ反应中心的过剩激发能Ex。在正常温度下,高羊茅吸收光能中的46.6%用于光化学反应,36.8%用于天线耗散,16.6%为PSII反应中心的过剩光能(表 3)。尽管高温处理使NPQ明显下降,但高温处理使天线耗散的能量的比例却略有上升,从对照的36.8%增加到39.3%;另外高温下PSII过剩光能的比例也显著上升,处理30 d后起增加到31.4%;用于光化学反应的部分则大量下降,说明高温处理降低光合作用CO2同化的电子需求。
|
|

|
|
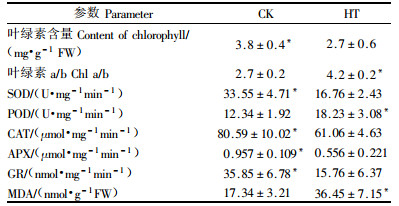
高温处理后,总Chl比对照降低29.3%,而Chla/b比对照上升35.7%。这说明Chlb比Chla下降得更快,而Chlb主要存在于2个光系统的天线色素中,这说明高温处理后叶片吸收和传递到PSⅡ反应中心的光能降低,这与前面提到的F′v/F′m等变化一致。另外,高温处理明显降低SOD,POD,CAT,APX和GR的活力,高温30 d后,SOD,POD,CAT,APX和GR活力分别比对照降低49.9%,47.7%,24.2%,41.9%和56.0%。但MDA含量却上升了52.4%。
3 讨论高温处理导致Pn的明显下降和光补偿点、CO2补偿点的上升, 这说明高温处理降低了高羊茅对光强和CO2的利用效率。根据Farquhar等(1980)提出的光合作用模型,高温处理30 d后,CE, Vc, max及Jmax都显著降低,而前二者直接取决于Rubisco羧化酶的活性变化, Jmax则取决于电子传递过程中同化力和RuBP再生系统中酶活性的高低(Long et al., 2003),因此高温胁迫明显降低Rubsico羧化酶的活性和RuBP再生速率,表现为CO2补偿点的上升。
高温胁迫30 d后,F′v/F′m, qP,ФPSⅡ和ETR等参数明显下降,说明长期高温使天线色素光能转换效率和PSⅡ反应中心开放部分的比例明显下降,用于CO2同化的光能减少,使过量的激发能在PSII反应中心的积累,从而降低光合电子传递能力,限制的RuBP再生能力。高温处理使叶绿素含量大量下降,Chla/b上升,说明Chlb的降解速度较快,从而使捕获和传递给PSII反应中心的光能减少。但高温处理明显使Fv/Fm减小,尽管传递给PSII的光能减少,但长期高温仍然使高羊茅遭受严重的光抑制,表现为光饱和点的明显下降。
植物在吸收的光能超过所利用的能量时,会启动非辐射能量的耗散途径来耗散过剩的激发能以保护光合机构免受伤害(Demmig-Adams et al., 1996)。本试验结果表明:长期高温处理后NPQ明显降低,长期高温胁迫下,高羊茅的能量耗散机制可能遭到伤害。qE是NPQ的主要部分,它依赖于跨类囊体膜的pH梯度(ΔpH),并受叶黄素循环等许多因素调控(Müller et al., 2001)。本研究中qE在长期高温下变化趋势与NPQ基本相同,表明NPQ的下降主要是由于qE的下降引起的。由于长期高温胁迫使电子传递降低,从而ΔpH不同有效的形成,使得能量耗散机制遭到伤害。qI也是NPQ的重要组成部分,主要光抑制程度相关,Rubisco活化酶的失活、D1蛋白的降解和PSII反应中心的失活可能在这种机制中起作用(Müller et al., 2001)。本研究结果表明高温胁迫使qI明显上升,说明长期高温使高羊茅受到明显的光抑制,这一结果与Fv/Fm的变化相吻合。
为了解高羊茅在长期高温胁迫下的光能利用情况,我们将植物吸收的光能分为3部分。结果表明:长期高温胁迫下PSII过剩光能Ex显著上升,用于光化学反应的能量则大量下降。Ex的增加导致光合电子传递到O2的相对比例增加,可形成更多的氧自由基、过氧化氢、羟基自由基等活性氧物质(徐红霞等,2005)。本研究中SOD,POD,CAT,APX和GR等抗氧化酶活力均大量下降,说明在长期的高温胁迫下,活性氧物质不能被抗氧化酶有效的清除掉。而MDA含量的大量上升说明活性氧已经积累到一定程度并诱发膜脂过氧化(Liu et al., 2000),这可能也是ΔpH不能有效形成的重要原因之一。
韩春梅, 张新全, 彭燕, 等. 2006. 18个高羊茅品种的耐热性比较研究. 草原与草坪, 1: 36-44. DOI:10.3969/j.issn.1009-5500.2006.01.008 |
何亚丽, 刘友良, 陈权, 等. 2002. 水杨酸和热锻炼诱导的高羊茅幼苗的耐热性与抗氧化的关系. 植物生理与分子生物学学报, 28: 89-95. |
徐红霞, 翁晓燕, 毛伟华, 等. 2005. 镉胁迫对水稻光合、叶绿素荧光特性和能量分配的影响. 中国水稻科学, 19: 338-342. DOI:10.3321/j.issn:1001-7216.2005.04.010 |
徐胜, 何兴元, 陈玮, 等. 2008. 热锻炼对高羊茅(Festuca arundinacea)和多年生黑麦草( Lolium perenne)抗高温能力的影响. 生态学报, 28: 162-171. DOI:10.3321/j.issn:1000-0933.2008.01.019 |
周嘉友, 汤承, 卢建霖, 等. 1998. 过渡带优质草坪建植与管理综合技术研究Ⅰ——坪床配方. 草业科学, 15(4): 44-48. |
Bilger W, Bjôrkman O. 1990. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynthesis Research, 25: 173-185. DOI:10.1007/BF00033159 |
Braber J M. 1980. Catalase and peroxidase in primary bean leaves during development and senescence. Z Pflanzenphysiology, 97: 135-144. DOI:10.1016/S0044-328X(80)80027-4 |
Cui L, Li J, Fan Y, et al. 2006. High temperature effects on photosynthesis, PSII functionality and antioxidant activity of two Festuca arundinacea cultivars with different heat susceptibility. Botanical Studies, 47: 61-69. |
Demmig-Adams B, Adams W W, Barker D H, et al. 1996. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation to of excess excitation. Physiol Plant, 98: 253-264. |
Foyer C H, Halliwell B. 1976. The presence of glutathione and glutathione reductase in chloroplasts:a proposed role in ascorbic acid metabolism. Planta, 133: 21-25. DOI:10.1007/BF00386001 |
Farquhar G D, von Caemmerer S, Berry J A. 1980. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta, 149: 78-90. DOI:10.1007/BF00386231 |
Genty B, Briantais J M, Baker N R. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta, 990: 87-92. DOI:10.1016/S0304-4165(89)80016-9 |
Giannopolitis C N, Ries S K. 1977. Superoxide dismutases:Ⅰ. occurrence in higher plants. Plant Physiology, 59: 309-314. DOI:10.1104/pp.59.2.309 |
Huang Z A, Jiang D A, Yang Y, et al. 2004. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence and antioxidant enzymes in leaves of rice plants. Photosynthetica, 42: 357-364. DOI:10.1023/B:PHOT.0000046153.08935.4c |
Jiang Y W, Huang B. 2001. Effects of calcium on antioxidant activities and water relations associated with heat tolerance in two cool-season grasses. Journal of Experimental Botany, 52: 341-349. DOI:10.1093/jexbot/52.355.341 |
Liu X Z, Huang B R. 2000. Heat stress injury in relation to membrane liqid peroxidation in creeping bentgrass. Crop Science, 40: 503-510. DOI:10.2135/cropsci2000.402503x |
Long S P, Bernacchi C J. 2003. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. Journal of Experimental Botany, 54: 2393-2401. DOI:10.1093/jxb/erg262 |
Maxwell K, Johnson G N. 2000. Chlorophyll fluorescence-a practical guide. Journal of Experimental Botany, 51: 659-668. DOI:10.1093/jexbot/51.345.659 |
Müller P, Li X P, Niyogi K K. 2001. Non-photochemical quenching. A response to excess light energy. Plant Physiology, 125: 1558-1566. DOI:10.1104/pp.125.4.1558 |
Nakano Y, Asda K. 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spiach chloroplasts. Plant Cell Physiology, 22: 867-880. |
Quick W P, Stitt M. 1989. An examination of factors contributing to non-photochemical quenching of chlorophyll fluorescence in barley leaves. Biochim Biophys Acta, 977: 287-296. DOI:10.1016/S0005-2728(89)80082-9 |
Thiele A, Winter K, Krause G H. 1997. Low inactivation of D1 protein of photosysem Ⅱ in young canopy leaves of Anacardium excelsum under light-light stress. Journal of Plant Physiology, 151: 286-292. DOI:10.1016/S0176-1617(97)80254-4 |
Zhao W Y, Xu S, Li J L, et al. 2008. Effects of foliar application of nitrogen on the photosyntheticperformance and growth of two fescue cultivars under heat stress. Biologia Plantrum, 52(1): 113-116. DOI:10.1007/s10535-008-0021-8 |