 2009, Vol. 45
2009, Vol. 45文章信息
- 李铁柱, 田大伦, 乌云塔娜, 谭晓风, 项文化, 闫文德.
- Li Tiezhu, Tian Dalun, Wuyun Tana, Tan Xiaofeng, Xiang Wenhua, Yan Wende
- 四倍体油茶的鉴定及变异
- Identification and Variation of Tetraploid Camellia oleifera
- 林业科学, 2009, 45(3): 150-154.
- Scientia Silvae Sinicae, 2009, 45(3): 150-154.
-
文章历史
- 收稿日期:2008-09-26
-
作者相关文章



2. 南方林业生态应用技术国家工程实验室 长沙 410004;
3. 经济林育种与栽培国家林业局重点实验室 长沙 410004;
4. 湖南会同杉木林生态系统国家野外科学观测研究站 会同 418300;
5. 城市森林生态湖南省重点实验室 长沙 410004
2. National Engineering Laboratory for Applied Technology of Forestry & Ecology in South China Changsha 410004;
3. Key Laboratory of Non-Wood Forest Product of State Forestry Administration Changsha 410004;
4. Huitong Ecological Station for Chinese Fir Plantation of Hunan Province Huitong 418300;
5. Key Laboratory of Urban Forest Ecology of Hunan Province Changsha 410004
油茶(Camellia oleifera )是我国南方主要的经济林木,与油棕(Elaeis gunieen)、油橄榄(Olea europaea)和椰子(Cocos nucifera)并称为世界四大木本食用油料树种。茶油不饱和脂肪酸含量达90%以上,以油酸和亚油酸为主,还含有少量的亚麻酸等高价不饱和脂肪酸,具有降低血清胆固醇含量,起到预防和治疗高血压和心血管疾病的作用(雷治国等,2003)。过去虽然通过常规选育方法培育了大量的油茶优良品种或优良无性系,但目前生产上仍缺乏油茶高产优良品种,导致油茶林经济效益较低,影响着油茶的大量栽培和进一步推广。
多倍体育种是有效的育种方法之一,在作物、园艺、林木育种中广泛应用(刘静等,2003;韩礼星等, 2003; 谷晓峰等, 2003; 王卓伟等, 2002; 张海洋等,2001; 李云等, 2001; 张海风等,2008)。染色体多倍化后由于基因剂量效应植物器官巨型化,茎粗、叶大而厚、花和果实增大增重,因此可以大幅度提高植物产量、品质、抗性等,因此多倍体的选育具有很好的生产应用价值(李凤兰等, 1994;Peloquin et al., 1992;Kaburu, 1992; Mashkina et al., 1998;Ramsey et al., 1998)。但是多倍体有可能导致育性及结实量降低(张海风等,2008)。油茶是以果实产量和产油率为目标的经济树种。从理论角度看,油茶多倍化后也许产油率会提高,但产籽率将会降低。在油茶栽培中有个特殊的问题,就是二次落果,即油茶除了正常的生理落果外,近成熟阶段还有一次大量落果的现象。这个现象有可能与油茶大量开花大量结果消耗了体内很多养分,后期养分跟不上有关,产量受到影响。如果四倍体油茶没有二倍体油茶结果量多,后期落果不严重,而且果实又大,这样或许能提高产量。本课题组以提高油茶产量为目标开展了普通油茶优良无性系多倍体诱导试验,本文报导了四倍体油茶的鉴定结果及其在形态、细胞学、分子水平上的变异。
1 材料与方法 1.1 植物材料秋水仙素诱导的植物材料是油茶-14、油茶-17、油茶-18三个普通油茶优良无性系,为湖南省林业科学院提供。
1.2 方法1) 秋水仙素处理 2005年2—4月,将油茶-14、油茶-17、油茶-18三个优良无性系种子播种到花盆中,待长出2个真叶后开始处理。处理方法:在顶芽上部缠绕少量棉花,分别滴2 g·L-1和3 g·L-1质量浓度的秋水仙素溶液至药棉浸透为止,2 h观察一次,始终保持湿润,持续滴药2 d。
2) 形态学观察和测定 2005,2006,2007连续3年在5—6月、9—10月观测形态性状,包括叶节间距、叶型、叶色、叶边缘锯齿、叶片大小等;采用LI23000A便携式叶面积分析仪测定植株中部成熟叶片的叶面积;用叶绿素测定仪测定叶片的叶绿素含量;叶片厚度采用石蜡切片法,显微镜下用显微测微尺量测;撕取叶片下表皮,在玻片上用1% I-KI染色后在40倍显微镜下用直线显微尺测量保卫细胞的长宽;于2008年9月采用美国产LI-COR公司生产的LI-6400便携式光合测量系统,测定秋水仙素诱导株和对照油茶的光合指标。数据统计分析采用Excel和SAS软件。
3) 染色体数目鉴定 2005年3—6月进行染色体数目分析。从0.2%~0.3%的秋水仙素诱导的油茶中筛选出长势好、形态特征明显发生变异的植株进行染色体数目分析,共68株。9:30取茎尖用对二氯苯饱和液处理后用卡诺氏液(95%乙醇:醋酸=3:1,体积比)固定12~24 h,5 mol·L-1 HCl酸解20~25 min,用40 g·L-1的硫酸铁铵进行媒染1 h,用0.5%苏木精染色4 h,经45%醋酸进行分色和软化后压片。将制好的片子放在显微镜载物台上,先用低倍镜(10×10)调焦观察,寻找具有分裂相的细胞后,再转到高倍镜(10×40)下观察,找到中期细胞后,最后转到油镜(10×100)下观察染色体数目。统计50个分裂中期的细胞染色体数,取85%以上细胞恒定一致的染色体数为试验植株的染色体数。
4) 总DNA含量测定及多态性分析 用购自北京天根生物有限公司的DNA提取试剂盒提取总DNA。DNA含量检测采用BeckmanDU-640型蛋白质核酸分析仪,测定230,260,280 nm处的紫外吸收光吸收值,根据260与280 nm的光吸收比值及260与230 nm的光吸收比值确定DNA的纯度,计算DNA的得率。电泳检测:0.8%琼脂糖凝胶,点样液5 μL,于BioRAD凝胶成像系统上观察电泳结果,拍照。总DNA的提取按购自北京天根生物有限公司的植物DNA提取试剂盒(离心柱型)说明书进行,略作改动。ISSR引物、线粒体DNA(mtDNA)和叶绿体DNA(cpDNA)遗传多样性检测引物共19组(表 1)。PCR反应用的2×Taq MasterMix购自北京天根生物有限公司;100 bp DNA Marker购自北京三博远志生物技术有限责任公司。PCR反应总体积为20 μL,含DNA template 1 μL,引物1 μL,2×Taq MasterMix 6.5 μL,SDW(ddH2O) 11.5 μL。PCR反应循环条件是94 ℃预变性2 min;94 ℃变性1 min,48 ℃复性40 s,72 ℃延伸2 min,40个循环;72 ℃延伸10 min;4 ℃保存。电泳凝胶浓度为1.2%。
|
|

所选出的68个植株中18个植株染色体数目为60条,为四倍体油茶(图 1A);44个植株的染色体数目不定,有些细胞中含有60条,而有些细胞中为30条,属于混合体;另外6个植株的染色体数目为30条,为二倍体(图 1B)。

|
图 1 四倍体油茶和二倍体油茶的染色体数目 Figure 1 Chromosome number in the diploid and tetraploid plants A:四倍体Tetraploid plants;B:二倍体Diploid plants. |
四倍体油茶苗冠幅大,植株粗壮,叶片增大且肥厚、变圆(图 2),叶色浓绿,叶间距和叶片锯齿间距大,茎粗,童期和花期比二倍体稍迟,这些表型符合多倍体的特征。

|
图 2 四倍体和二倍体植株的叶片 Figure 2 The leaf of diploid and tetraploid plants A, C:二倍体Diploid plants;B, D:四倍体Tetraploid plants. |
从表 2可以看出,四倍体与二倍体植株的叶面积、叶长、叶宽、叶厚之间存在显著差异,叶形指数和叶间距之间存在极显著差异,叶片锯齿数差异不明显。四倍体油茶与二倍体油茶相比,其叶面积、叶长、叶宽、叶厚、叶间距分别增加9.8%, 38.5%, 25%, 48.25%, 67%,而叶形指数减少38.1%。
|
|
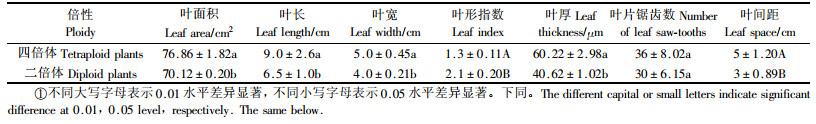
对四倍体油茶和二倍体油茶叶绿素含量、保卫细胞的特点进行了观察和分析。结果表明,四倍体油茶和二倍体油茶叶绿素含量存在极显著的差异(表 3),四倍体叶绿素含量比二倍体增加47.87%。保卫细胞长度是区别于多倍体和二倍体的重要标志,从表 3中可看出,四倍体油茶叶片保卫细胞的长、宽明显大于二倍体油茶,分别比二倍体增加48.15%和37.26%。
|
|

从四倍体油茶和二倍体油茶的光响应曲线图(图 3)可看出,四倍体油茶叶净光合速率比二倍体油茶高。

|
图 3 二倍体油茶和四倍体油茶光响应曲线 Figure 3 Light response curves of diploid and tetraploid plants |
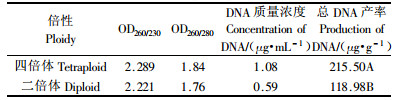
利用BeckmanDU-640型蛋白质核酸分析仪测定四倍体油茶和二倍体油茶DNA含量(表 4)。从表 4可看出,100 mg叶片中的总DNA得率四倍体油茶约215.50 μg,而二倍体油茶约118.98 μg,四倍体的总DNA得率是二倍体油茶的近2倍。从四倍体和二倍体油茶的基因组电泳图(图 4A)可看出,四倍体油茶总DNA条带比二倍体油茶更亮。
|
|

|
图 4 四倍体和二倍体油茶的DNA遗传多态性 Figure 4 The electrophoresis patterns of polymorphic in the diploid and tetraploid plants 1:二倍体Diploid plants; 2:四倍体Tetraploid plants;A:基因组DNA Genome DNA; B, C, D, E, F:mtDNA和cpDNA引物PCRThe PCR of mtDNA and cpDNA; H, I, J, K:ISSR引物PCR扩增The PCR of ISSR. |
叶绿体DNA和线粒体DNA具有母性遗传特点,为多倍体鉴定提供了依据。利用线粒体DNA和叶绿体DNA的8对引物对四倍体油茶和二倍体油茶总DNA进行PCR扩增。结果表明,8对引物在四倍体油茶和二倍体油茶中扩增出11个条带,所检测的11基因位点均未出现遗传多态性,这说明所获得的四倍体油茶和对照二倍体之间叶绿体DNA和线粒体DNA遗传基因是一致的,同时说明了四倍体油茶是对照二倍体油茶染色体加倍而来的。但ISSR分析表明,11条引物共扩增出37个基因位点,其中四倍体油茶和二倍体油茶在19基因位点上存在遗传差异,多态性位点比率达51.35%,这说明了染色体加倍后四倍体油茶在染色体结构上有可能发生了变化(王卓伟等,2002)。
3 讨论诱变育种是非常有效的常规育种方法,诱变率可达1/1 000。其中秋水仙素诱导产生多倍体在很多植物中应用过(王柏龄等,1999;刘静等,2003;韩礼星等, 2003; 谷晓峰等, 2003; 王卓伟等, 2002; 张海洋等, 2001; 李云等, 2001; 康向阳等, 1999;张海风等,2008)。作者所在课题组对普通油茶优良无性系进行多倍体诱导试验,获得了多株突变体。这些突变体有的产生染色体加倍(叶、花、果实增大),有的只发生形态变化(整株出现一侧长叶另一侧不长叶),也有的出现早花早果,这些突变体为油茶优良品种选育、遗传改良提供了良好的物质基础。本研究选用的试验材料就是从众多突变株中筛选出的具有多倍体特征的植株,对其进行了多倍体鉴定。对照普通油茶优良无性系为二倍体,染色体数目为2n=2x=30,而诱变株染色体数目为2n=4x=60,为四倍体。
四倍体油茶和二倍体油茶在形态特征、保卫细胞特征(刘静等,2003;张海洋等, 2001;张海风等,2008)、叶绿素含量、光合作用、DNA含量上均存在显著或极显著差异。在形态方面四倍体油茶叶片厚而大、叶色深绿、节间长、茎粗。叶片保卫细胞长而宽,叶绿素含量增加,净光合速率也提高,DNA含量是二倍体油茶的近2倍。多倍体一般表现为生长快,抗逆性强(张海风等,2008),花、果实等生殖器官比二倍体增大。多倍体在遗传学效应上确实具有提高生长速度、抗逆性、花和果实大小等特点,但有可能导致育性降低以及结实量降低。在油茶栽培中一个特别的问题,就是二次落果,也就是油茶除了正常的生理落果外,近成熟阶段还有一次大量落果的现象。这个现象有可能与油茶大量开花大量结果消耗了体内很多养分,后期养分跟不上有关,产量受到很大影响。如果四倍体油茶没有二倍体油茶结果量多,后期落果不严重,而且果实又大,这样或许能提高产量,当然这需要进一步的试验研究。
谷晓峰, 罗正荣. 2003. 秋水仙素处理'罗田甜柿'获得12倍体再生植株. 园艺学报, 30(3): 325-327. DOI:10.3321/j.issn:0513-353X.2003.03.019 |
韩礼星, 赵改荣, 李玉红. 2003. 猕猴桃多倍体诱导研究. 果树科学, 15(3): 273-276. |
康向阳, 朱之悌, 林惠斌. 1999. 杨树花粉染色体加倍有效处理时期的研究. 林业科学, 35(4): 21-24. DOI:10.3321/j.issn:1001-7488.1999.04.004 |
雷治国, 黄永芳, 何会蓉. 2003. 油茶及种质资源研究进展. 经济林研究, 21(4): 123-125. DOI:10.3969/j.issn.1003-8981.2003.04.041 |
李凤兰, 张志毅. 1994. 白杨染色体加倍技术研究及三倍体育种. 北京林业大学学报, 16(2): 15-18. |
李云, 朱之悌, 田砚亭. 2001. 秋水仙碱处理白杨雌花芽培育三倍体植株的研究. 林业科学, 37(5): 68-74. DOI:10.3321/j.issn:1001-7488.2001.05.012 |
刘静, 丁兆堂, 赵进红, 等. 2003. 茶树多倍体诱变研究初报. 山东农业大学学报, 34(4): 475-478. DOI:10.3969/j.issn.1000-2324.2003.04.004 |
王柏龄, 万辉, 李烈国. 1999. 秋水仙素诱变茶树短穗腋芽的试验. 蚕桑茶叶通讯, 12(1): 7-9. |
王卓伟, 余茂德, 鲁成. 2002. 桑树二倍体及人工诱导的同源四倍体遗传差异的AFLP分析. 植物学通报, 19(2): 194-200. DOI:10.3969/j.issn.1674-3466.2002.02.009 |
张海风, 郭宝林, 张成合, 等. 2008. 杜仲多倍体的诱导与鉴定. 园艺学报, 35(7): 1047-1052. DOI:10.3321/j.issn:0513-353X.2008.07.016 |
张海洋, 卫双玲, 卫文星. 2001. 芝麻同源四倍体的诱发与鉴定. 华北农学报, 16(2): 12-15. DOI:10.3321/j.issn:1000-7091.2001.02.003 |
Kaburu H. 1992. Fertility of doubling monoploids of Solanum phureja. American Potato Journa, 69: 447-459. DOI:10.1007/BF02852295 |
Mashkina O S, Burdaeva L M, Belozerova M, et al. 1998. Method of obtaining diploid pollen of woody species. Lesovedenie, 34(1): 19-25. |
Peloquin S J, Ortiz R, Stalker H T, et al. 1992. Plant breeding in the 1990s. Wallingford, PA: CAB International, 485-507.
|
Ramsey J, Schemske D W. 1998. Pathway, mechanisms, and rates on polyploid formation in flowering plant. Annual Review of Ecology and Systematics, 29(2): 467-501. |