 2009, Vol. 45
2009, Vol. 45文章信息
- 陈美玲, 上官周平.
- Chen Meiling, Shangguan Zhouping
- 黄土高原子午岭林区6个典型群落优势种的热值和养分特征
- Caloric Value and Nutrient Characteristics of Dominant Plant Species of Six Typical Vegetation Communities in Ziwuling Forest Area of the Loess Plateau
- 林业科学, 2009, 45(3): 140-144.
- Scientia Silvae Sinicae, 2009, 45(3): 140-144.
-
文章历史
- 收稿日期:2007-05-08
- 修回日期:2008-12-25
-
作者相关文章



2. 中国科学院水土保持研究所 杨凌 712100
2. Institute of Soil and Water Conservation, Chinese Academy of Sciences Yangling 712100
能量流动、物质循环和信息传递共同构成了生态系统三大功能。能量是维持生态系统功能与过程的动力,Jordan(1971)认为能量比干物质更能反映出群落对自然资源(特别是太阳能)的利用情况。Long(1934)率先用热值来表示植物所含能量的多少,此后关于植物热值的研究工作逐渐展开。我国的植物热值研究始于20世纪80年代(杨福囤等,1983;祖元刚等,1986),主要集中在海南(林益明等,2000)、福建(林益明等,2004)、广东(任海等,1999)和浙江(郝朝运等,2006)等热带亚热带植物群落以及内蒙古羊草草原(鲍雅静等,2003),但有关黄土高原地区的植被热值研究尚未见报道。子午岭林区是黄土高原主要的天然次生林区,其植被主要是在弃耕地基础上逐渐恢复起来的(邹厚远等,2002)。由于自然因素与人为因素的影响,子午岭的植被在恢复时间上存在较大差异,形成了处于不同演替阶段的各种植被群落类型。本研究选取的6个代表性植被群落分别是沙棘(Hippophae rhamnoides)群落、山杨(Populus davidiana)群落、白桦(Betula platyphylla)群落、油松(Pinus tabulaeformis)群落、柴松(Pinus tubuleformis f. shekansis)群落和辽东栎(Quercus liaotungensis)群落,群落年龄分别为15,39,41,38,43和42 a,这6个群落可以通过时空互代的方法看作同一森林群落演替过程中的6个不同阶段。对于不同演替阶段的植被类型来说,它们能够形成特定的耗散结构类型,体现出不同阶段的能量结构特征,因此可利用能量当量来研究生态系统的进程和格局变化(乔秀娟等,2007)。本研究的目的就是揭示子午岭林区植被演替不同阶段群落类型优势树种及凋落物的热值特征,同时从热值角度对子午岭林区特有种柴松群落在森林演替中的地位进行初步探讨,为子午岭林区森林植被的合理管理和保护、人工植被自然化抚育与营造提供理论依据。
1 研究区概况研究区位于黄土高原子午岭林区北部甘肃省合水县连家砭林场(35°03′—36°37′ N,108°10′—109°08′ E),该区属黄土高原丘陵沟壑区,海拔1 200~1 600 m,相对高差100~300 m,为半干旱季风气候,梁峁顶部浑圆平缓,斜倾3°~5°,沟坡上斜下陡,变化于10°~35°之间。该区25年平均气温7.4 ℃,年均降水量587.6 mm,年平均相对湿度63%~68%,≥10 ℃年积温2 671.0 ℃, 干燥度0.97,阴阳坡水热条件变化较大但无气候的垂直带状变化。土壤为原生(山坡)或次生(沟谷)黄土,厚度一般为50~100 m,其下为厚80~100 m的红土,以石灰性褐土为主,其次为粗骨褐色土(邹厚远等,2002)。
2 研究方法 2.1 样品采集于2005年植物生长季7月中旬,选取研究区处于不同演替阶段的代表性群落设立标准样地(20 m × 20 m),尽量保持海拔和坡向的一致,以减小环境条件的差异对试验结果的影响。在标准样地内进行每木检尺,选择与标准木相近的优势木5株,分器官采样,叶片采成熟健康叶,枝和根按不同发育程度采样,保证粗枝和细枝,粗根和细根比例均衡,皮和干在树木胸径处(距地面1.3 m)采样,分别标记、混匀后装入自封袋,并迅速带回试验基地。于105 ℃下杀青10 min,70 ℃下烘干至恒质量,供养分和热值的测定。
用遮阳网采集新鲜凋落物。于7月初在样地范围内挂1 m × 1 m的遮阳网,分6个方位,每个方位2张(间隔2~3 m),夹角60°,离地面20 cm,7月中旬和下旬分别收集1次。凋落物残体分6个方位采集,每个方位采集2簇,在20 m × 20 m样地范围内,将采集的样品仔细混匀,装入布袋带回试验基地。70 ℃下烘干至恒质量后将样品磨细分别过0.154,0.25和0.5 mm筛后装入玻璃瓶,放入干燥器供养分、热值和灰分测定。在7月下旬采集遮阳网下1 m × 1 m的地被物为凋落物残体样品。
2.2 有机碳、全氮和全磷的测定采用外加热、重铬酸钾容量法测定有机碳含量(mg·g-1)。全氮测定用凯氏定氮法(mg·g-1)。用钼蓝比色法测定全磷含量(mg·g-1)。
2.3 热值与灰分的测定热值用热量计法测定,仪器采用XRY-1A型数显氧弹式热量计(上海)。用空调控温在(22±2) ℃,室内杜绝热源和空气对流,每份样品重复3~5次。分别测定样品的干质量热值和去灰分热值,试验前用国家二级热量标准物质苯甲酸对仪器进行标定。灰分含量的测定用干灰化法(Paine,1971),即样品在马福炉500 ℃下灰化4 h后测定其灰分含量(mg·g-1)。活立木热值取植物根、皮、干、枝和叶的平均值。
2.4 数据分析用SAS(8.0)统计分析软件对数据进行相关分析与One-way ANOVA方差分析,并用LSD法进行多重比较。
3 结果与讨论 3.1 优势树种的干质量热值和去灰分热值在植被演替的不同阶段,随着优势植物种类的相互替代,群落光照和土壤理化性质以及群落环境条件的变化导致群落优势树种热值结构的变化。6个优势种各器官干质量热值为(16.974±0.100) kJ·g-1~(21.500±0.268) kJ·g-1,辽东栎枝最低,柴松叶最高(表 1)。辽东栎各器官干质量热值均显著低于其他优势种(F= 3.42,P<0.05),平均干质量热值从高到低的顺序为油松>柴松>白桦>山杨>沙棘>辽东栎。
6个优势种去灰分热值之间的差异较小,介于(17.220±0.185) kJ·g-1~(22.427±0.787) kJ·g-1之间,辽东栎枝最低,山杨树皮最高,辽东栎各器官的干质量热值均显著低于其他优势种(F= 3.66,P<0.05)。去灰分热值加权平均值从高到低排列为山杨>油松>柴松>白桦>沙棘>辽东栎。辽东栎的平均干质量热值和平均去灰分热值均为最低,先锋植被沙棘的干质量热值和去灰分热值也低于处于演替中期的山杨、白桦和油松(表 1),表明随着群落的正向演替,优势树种的热值呈现先增加后降低的趋势。乔秀娟等(2007)研究了西双版纳不同林龄次生植物群落优势树种的热值,也得出各群落优势树种平均热值前期上升,后期略有下降的结论。由于热值反映的是绿色植物在光合作用中转化光能的能力(Pitelka,1977;郑帷婕等,2007),这种先增加后降低的热值变化趋势可能表明了优势植被对光能的利用效率随着演替的进展呈现首先提高然后降低的趋势。柴松各器官的干质量热值和去灰分热值以及加权平均值均较高,说明柴松具有较高的光能利用效率,柴松群落可能处于演替中期或后期,属于亚顶级群落。
|
|

6种优势植物叶、枝、皮、干和根的平均干质量热值分别为(19.870±1.075) kJ·g-1,(19.114±1.161) kJ·g-1,(19.481±0.722) kJ·g-1,(18.984±0.366) kJ·g-1和(18.883±0.555) kJ·g-1,存在着叶>皮>枝>干>根的规律,t检验表明叶片干质量热值显著大于根(t = 2.91,P<0.05),其余各器官之间的差异均不显著。6种优势植物叶、枝、皮、干和根的平均去灰分热值分别为(20.656±1.069) kJ·g-1,(19.613±1.269) kJ·g-1,(20.415±1.285) kJ·g-1,(19.062±0.269) kJ·g-1和(19.604±0.825) kJ·g-1,叶片仍然是含热值最高的器官,变化规律与干质量热值略有不同,为叶>皮>枝>根>干,灰分影响了根和干的去灰分热值排序。叶片去灰分热值显著大于树干(t = 2.87,P<0.05),其他器官之间去灰分热值没有明显差异。由于枝、皮、干和根等器官含有较多的木质素,木质素的热值较低,而叶片含有丰富的叶绿体,叶绿体主要由蛋白质和脂肪等构成(郑帷婕等,2007),蛋白质和脂肪的热值较高(祖元刚,1990),这可能是叶的干质量热值和去灰分热值都显著高于其他器官的原因。
3.2 不同群落活立木、新鲜凋落物和凋落物残体的热值与养分含量由于不同演替阶段群落存在微生境差异,有机质的积累和分解过程也不同(表 2)。在子午岭林区植被演替中沙棘是演替早期种,山杨、白桦是演替中期种,油松是演替后期种,辽东栎是子午岭林区的顶级种(邹厚远等,2002)。除山杨群落外,各群落有机碳含量均为活立木>新鲜凋落物>凋落物残体,全氮和全磷含量没有明显变化规律。演替早期和中期群落能量流动波动较大,后期群落能量流动趋于稳定,植物生长季节的新鲜凋落物主要成分为叶,小枝和树皮等只占很小比例,由于叶片中的有机物质以蛋白质和脂肪为主,并积累了大量的光合产物,叶片在凋落以前主要发生养分元素的转移,保留了有机物质,使得尚未开始分解的新鲜凋落物还有较高的能量储备,而活立木中根、干、枝等所占生物量比重大,且主要由木质素和纤维素等低热值的有机物质构成,这可能是活立木的加权平均热值总体低于新鲜凋落物热值的原因。凋落物残体的营养物质和高能有机物质率先被土壤微生物分解和释放,随着分解的进行,凋落物残体中低热值有机物质所占比例增加,这可能是3个组分中凋落物残体的干质量热值和去灰分热值均最低的原因。
|
|

凋落物残体的分解是能量和物质释放的重要途径,土壤中的微生物在其中发挥了极为重要的作用。对活立木、新鲜凋落物与凋落物残体3者热值显著性分析表明,凋落物残体干质量热值与活立木和新鲜凋落物的干质量热值均有显著差异,3个组分去灰分热值之间差异没有达到显著水平。在比较不同植物热值时采用去灰分热值更为合理,这样可以消除灰分含量的不同带来的影响(Reiners,1972)。活立木、新鲜凋落物与凋落物残体去灰分热值的比率分别表现为白桦群落1.098:1.082:1、油松群落1.352:1.386:1、柴松群落1.210:1.262:1、辽东栎群落1.307:1.301:1,沙剌群落的活立木与凋落物残质的比率为1.068:1,山杨群落的活立木与新鲜凋落物的比率为1.006:1。油松和辽东栎群落具有较高的能量回归比例,柴松群落能量回归比例大于沙棘、山杨和白桦群落,介于油松和辽东栎群落之间,说明柴松群落在子午岭森林演替进程中接近演替后期。
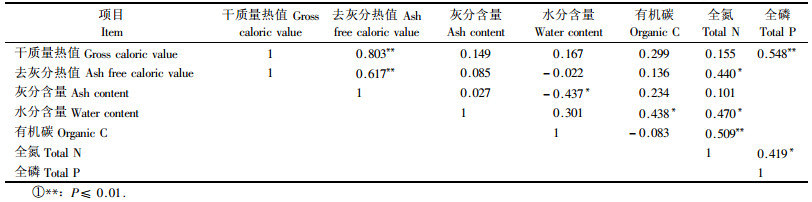
3.3 植物各器官热值与元素含量的相关性在植物生长状况良好的情况下,有机碳和全氮含量对热值不存在显著影响,全磷含量与干质量热值和去灰分热值具有极显著和显著正相关关系。磷是细胞核的主要成分,是植物体内各种代谢活动以及生殖生长非常重要的元素,Pitelka(1977)认为植物的器官拥有高热值,所以植物热值会随着全磷含量的增加而呈不同程度地增长。去灰分热值与灰分含量有极显著的正相关关系,干质量热值与灰分含量相关性不显著。干质量热值与去灰分热值极显著相关。去灰分热值是指植物体有机物质的热值,脂肪、蛋白质、碳水化合物、木质素、纤维素、淀粉等共同构成了植物有机质,它们的热值差异很大,祖元刚(1990)曾对它们的热值做过详细的研究,其中脂肪的热值最高39.54 kJ·g-1,淀粉的热值较低15.96 kJ·g-1,各种有机物质的含量是影响热值高低的重要因素(表 3)。
|
|
子午岭是黄土高原至今保存最为完整的天然次生林区,特殊的地理位置使得子午岭森林植被在过去150多年的恢复过程中形成了其独特的生态系统,具有在空间上完整的演替序列。随着植被群落正向演替,子午岭林区优势植被的干质量热值和去灰分热值呈现先逐渐提高然后降低的趋势,柴松具有较高的干质量热值和去灰分热值,表明柴松群落可能为处于演替中期或后期的亚顶级群落。
叶片具有明显高于其他器官的干质量热值和去灰分热值(P<0.05),其他器官之间干质量热值和去灰分热值的差异不明显。全磷在植被热值组成中占有重要地位,并与干质量热值和去灰分热值呈显著正相关。
在植物体逐渐向土壤回归的过程中,蛋白质和脂肪等高能物质被率先分解回归,植物热值逐渐降低,且物质组成逐渐变得较为简单,影响热值的因子开始减少。凋落物残体具有较高的能量回归速率,各群落凋落物残体的热值总体低于活立木和新鲜凋落物。
鲍雅静, 李政海. 2003. 内蒙古羊草草原群落主要植物的热值动态. 生态学报, 23(3): 606-613. DOI:10.3321/j.issn:1000-0933.2003.03.025 |
郝朝运, 刘鹏. 2006. 浙江北山七子花群落主要植物叶热值. 生态学报, 26(6): 1709-1718. DOI:10.3321/j.issn:1000-0933.2006.06.010 |
林益明, 林鹏, 王通. 2000. 几种红树植物木材热值和灰分含量的研究. 应用生态学报, 11(2): 181-184. DOI:10.3321/j.issn:1001-9332.2000.02.006 |
林益明, 郭启荣, 叶功富, 等. 2004. 福建东山几种木麻黄的物质与能量特征. 生态学报, 24(10): 2217-2224. DOI:10.3321/j.issn:1000-0933.2004.10.019 |
乔秀娟, 曹敏, 林华. 2007. 西双版纳不同林龄次生植物群落优势树种的热值. 植物生态学报, 31(2): 326-332. DOI:10.3321/j.issn:1005-264X.2007.02.017 |
任海, 彭少麟, 刘鸿先, 等. 1999. 鼎湖山植物群落及其主要植物的热值研究. 植物生态学报, 23(2): 148-154. DOI:10.3321/j.issn:1005-264X.1999.02.006 |
杨福囤, 何海菊. 1983. 高寒草甸地区常见植物热值的初步研究. 植物生态学与地植物学丛刊, 7(4): 280-288. |
张东来, 毛子军, 张玲, 等. 2006. 森林凋落物分解过程中酶活性研究进展. 林业科学, 42(11): 105-109. |
郑帷婕, 包维楷, 辜彬, 等. 2007. 陆生高等植物碳含量及其特点. 生态学杂志, 26(3): 307-313. |
邹厚远, 刘国彬, 王涵生. 2002. 子午岭林区北部近50年植被的变化发展. 西北植物学报, 22(1): 1-8. |
祖元刚, 张宏一. 1986. 植物热值测定的若干技术问题. 生态学杂志, 5(4): 53-56. |
祖元刚. 1990. 能量生态学引论. 长春: 吉林科学技术出版社.
|
Andersson M, Kj¢ller A, Struwe S. 2004. Microbial enzyme activities in leaf litter, humus and mineral soil layers of European forests. Soil Biology and Biochemistry, 36(10): 1527-1537. DOI:10.1016/j.soilbio.2004.07.018 |
Jordan C F. 1971. Productivity of a tropical forest and its relation to a world pattern of energy storage. Journal of Ecology, 59: 127-142. DOI:10.2307/2258457 |
Long F L. 1934. Application of calorimetric methods to ecological research. Plant Physiology, 9(2): 323-327. DOI:10.1104/pp.9.2.323 |
Paine R T. 1971. The measurement and application of the calorie to ecological problem. Annual Review of Ecology and Systematics, 2: 145-164. DOI:10.1146/annurev.es.02.110171.001045 |
Pitelka L F. 1977. Energy allocations in annual and perennial lupines (Lupinus:leguminosae). Ecology, 58(5): 1055-1065. DOI:10.2307/1936925 |
Reiners W A. 1972. Comparison of oxygen-bomb combustion with standard ignition techniques for determining total ash. Ecology, 53(1): 32-136. |
Sariaslani F S, Dalton H. 1989. Microbial enzymes for oxidation of organic molecules. Critical Reviews in Biotechnology, 9(3): 171-257. DOI:10.3109/07388558909036736 |