 2009, Vol. 45
2009, Vol. 45文章信息
- 曹志勇, 邵芬娟, 刘亚娟, 范俊玲, 魏学智.
- Cao Zhiyong, Shao Fenjuan, Liu Yajuan, Fan Junling, Wei Xuezhi
- 杜松小孢子的发生和雄配子体的发育
- Microsporogenesis and Male Gametophyte Development of Juniperus rigida
- 林业科学, 2009, 45(3): 74-78.
- Scientia Silvae Sinicae, 2009, 45(3): 74-78.
-
文章历史
- 收稿日期:2008-02-27
-
作者相关文章



杜松(Juniperus rigida)也称棒松和崩松,为柏科刺柏属(Juniperus)常绿小乔木。主要生长于我国黑龙江、吉林、辽宁、内蒙古、山西等地,日本、朝鲜也有分布。海拔1 000~2 200 m的半阳坡,耐干旱瘠薄的土壤。其树冠塔形或圆柱形,可作园林绿化树种;其材质坚硬致密,可供工艺、雕刻、盆景等用(中国植物志,2000);种子可榨油,也可供药用,有发汗、镇痛和利尿等功效(高艳平等,2005)。
由于杜松树形美观,极具观赏价值,已有许多学者对其研究但多集中在栽培育种(邹国林等,1996;曹泽猷,1993;卫金圈,2001;孙殿宝等,2005)、病原菌防治(张剑斌等,2000;董希文等,2002)和化学成分分析(张少贞等,1996;张国秀等,1994),有关杜松的胚胎学资料,迄今尚未见报道。植物生殖是物种繁殖和生存的保证,鉴于杜松的观赏、绿化、药用和防风固沙等作用,有必要对其生殖生物学问题进行深入的研究,旨在为杜松生殖生物学提供理论基础并从生殖过程角度探讨其与环境的适应性。本文通过石蜡制片对杜松小孢子发生和雄配子体的发育进行较为细致的观察和报道。
1 材料和方法杜松采自山西灵石石膏山森林自然保护区海拔1 000~1 300 m范围内。从2006年3月至2007年10月,每10 d采集1次,散粉前后每天采集,固定于FAA(70%乙醇:乙酸:福尔马林=18:1:1)中。常规石蜡法制片,切片厚度5~8 μm,铁矾-苏木精染色,中性树胶封片,显微镜观察并摄影(李正理,1996)。
2 观察结果 2.1 小孢子叶球杜松为雌雄异株植物,6~7年的雄株便可形成小孢子叶球。其小孢子叶球着生于雄株2年生枝叶腋中,单生,卵圆形,纵径为4~5 mm(图版Ⅰ-1)。小孢子叶球由4~5轮小孢子叶组成,每轮3枚,其基部有2片苞片和2~3轮鳞片包裹(图版Ⅰ-2)。3月初,幼嫩的小孢子叶球开始发生,呈圆锥形,乳黄色,由鳞叶紧紧包裹。3月中下旬,小孢子叶球继续生长露出鳞叶外,浅绿色,呈球形。4月中旬,小孢子叶球由浅绿色变成黄褐色,体积明显增大。4月下旬至5月上旬,小孢子叶球由黄褐色经淡黄色慢慢变成乳白色,小孢子叶和小孢子囊柄伸长,使紧密的小孢子叶变得松散,小孢子囊完全暴露在空气中,以利于传粉。
图版说明 1.雄株枝条;2.小孢子叶球(1.小孢子叶;2.鳞片;3.苞片);3.小孢子叶基部远轴面和近轴面开始分化出小孢子囊,×100;4.小孢子囊,示小孢子囊壁的表皮,中层,绒毡层,×400;5.表皮细胞内柱状结构,×400;6.退化的中层,×400;7.双核绒毡层细胞横切面观,×400;8.退化的绒毡层,×400;9.小孢子叶表皮下细胞分裂活跃,×400;10.孢原细胞,×400;11.初生造孢细胞,×400;12.小孢子囊和次生造孢细胞,×400;13.小孢子囊和小孢子母细胞,×400;14.部分小孢子母细胞退化,在小孢子囊内形成空腔,×400;15.游离的小孢子母细胞,×400;16~20.小孢子母细胞减数分裂前期Ⅰ的细线期、偶线期、粗线期、双线期和终变期,×1 000;21~23.小孢子母细胞减数分裂Ⅰ的中期、后期和末期,×1 000;24.四面体形末期Ⅱ,×1 000;25.左右对称形末期Ⅱ,×1 000;26.四分体小孢子壁同时形成,×1 000;27.小孢子母细胞减数分裂,×1 000;28.四面体形和左右对称形四分体,×400;29.单核小孢子,×1 000;30.单核靠边期,×1 000;31.成熟花粉粒,×1 000;32~33.胚珠中的小孢子胚性细胞,×100(×1 000);34~35.胚性细胞分裂形成生殖细胞和管细胞,×100(×1 000);36~37.生殖细胞分裂形成精原细胞和不育细胞,×100(×1 000);38~39.精原细胞分裂形成2个大小相似的精子细胞,×100(×1 000).MMC:小孢子母细胞;ST:造孢组织;Ep:表皮;Ml:中层;T:绒毡层。
Explanation of plate 1.A Branch of male;2.The microstrobilus(1.The microsporophylls; 2.The scale; 3. The bract);3.The microsporophylls bear their microsporangia on the abaxial and adaxial surface,×100;4.Transverse section of the microsporangium,showing epidermis,middle layers,tapetum,×400;5. The column structure in the epidermis cells,×400;6. The degenerated Ml cells,×400;7. Transverse section of the tapetal cells consisting of two nuclei,×400;8. The degenerated T cells,×400;9.The active cell division under the microsporophylls epidermis,×400;10.The archesporial cell,×400;11.The primary sporogenous cells,×400;12.The microsporangium and the secondary sporogenous cells,×400;13.The microsporangium and MMC,×400;14.Part of MMC degenerate,resulting in the formation of a large cavity in the microsporangium,×400;15.Free MMC,×400;16~20.ProphaseⅠof MMC meiosis,showing leptotene,amphitene,pachytene,diplotene and diakineses, ×1 000; 21~23.The stages of MMC meiosisⅠ,showing metaphasel,anaphase and telophase; 24. Tetrahedral telophaseⅡ, ×1 000; 25. Isobilateral telophaseⅡ, ×1 000; 26. The simultaneity of tetrad microspore wall, ×1 000; 27. The meiosis of MMC, ×1 000; 28.Tetrahedral and isobilateral tetrad, ×400; 29.The uninucleate microspore, ×1 000; 30. The vacuolated period of microspores, ×1 000; 31.Mature one-cell pollen, ×1 000; 32~33. The microspore embryonic cell in the ovule, ×100(×1 000); 34~35. Formation of generative cell and tube nucleus, ×100(×1 000); 36~37.Formation of spermatogenous cell and sterile cell, ×100(×1 000); 38~39. Formation of two sperm cells×100, (×1 000).
MMC:Microspore mother cells;ST:Sporogenous tissue;Ep:Epidermis;Ml:Middle layer;T:Tapetum.

|
图版Ⅰ Plate Ⅰ |
成熟的小孢子叶远轴面近基部着生2~4个小孢子囊,无柄;小孢子叶近轴面生1~8个小孢子囊,这些小孢子囊簇生在共同的孢子囊柄上。小孢子叶近轴面的小孢子囊数目,从最外一轮小孢子叶,至最内一轮小孢子叶,依次递减。在解剖镜下观察,最外一轮小孢子叶近轴面的小孢子囊,大部分直接着生在孢子囊柄上,小部分2~3个着生在次级小孢子叶远轴面,而次级小孢子叶着生在孢子囊柄上(图 1)。

|
图 1 成熟小孢子叶球示意图 Figure 1 The sketch map of mature microstrobilus 1.小孢子囊柄The microsporangia pedicle;2.中轴Axis;3.小孢子叶Microsporophylls;4.次级小孢子叶Second microsporophylls;5.近轴面小孢子囊Adaxial microsporangia;6.远轴面小孢子囊Microsporangia;7.鳞片Scale;8.苞片Bract. |
3月初到3月中旬,小孢子叶分化期结束,小孢子叶基部远轴面和小孢子叶叶腋处的细胞分裂加快,开始分化出小孢子囊(图版Ⅰ-3)。杜松小孢子囊一般由3层细胞组成,包括表皮、中层和绒毡层(图版Ⅰ-4)。在小孢子发生和雄配子体发育过程中,表皮细胞不断进行垂周分裂,增加细胞数目,以适应内部组织的迅速生长;其径向壁和内切向壁也开始次生加厚,高度木质化(图版Ⅰ-5)。花粉粒成熟时,表皮细胞内出现透明柱状结构,与表皮细胞壁连为一体,成为表皮细胞的支架(图版Ⅰ-5)。中层紧贴表皮细胞,细胞质稀少,随着内部造胞细胞数目的增多和体积的增大,被挤压而逐渐变的扁平(图版Ⅰ-4,12,13)。在以后小孢子母细胞发育过程中,中层细胞被挤压成条形,逐渐被吸收和解体,到小孢子形成时,中层细胞退化仅存残迹(图版Ⅰ-6)。中层的内方是绒毡层细胞,随着造孢细胞的发育,绒毡层细胞的细胞质变浓,细胞核大,随着小孢子囊内部压力的不断增大,绒毡层细胞变为扁平形状(图版Ⅰ-6,12),至造孢组织形成时,绒毡层细胞进行分裂形成双核细胞,从横切面上看,细胞内含有几个小液泡(图版Ⅰ-7)。随着小孢子母细胞的分裂,绒毡层细胞逐渐解体,至小孢子液泡化时,绒毡层细胞仅存残迹(图版Ⅰ-8),当花粉粒发育成熟时,小孢子囊壁只剩下高度木质化的表皮层(图版Ⅰ-5)。
2.3 小孢子的发生3月中旬,在杜松小孢子叶基部远轴面和小孢子叶近轴面,表皮下出现一群分裂活跃的细胞,表皮细胞也进行平周分裂,逐渐分化出小孢子囊(图版Ⅰ-9)。位于小孢子囊中央的细胞体积增大、细胞核大、细胞质浓稠,这些细胞即为孢原细胞(图版Ⅰ-10)。孢原细胞进行有丝分裂,形成初生造孢细胞(图版Ⅰ-11)。初生造孢细胞继续进行有丝分裂产生次生造孢细胞。3月下旬,由于次生造孢细胞的不断分裂,小孢子囊内充满密集的造孢细胞,形成造孢组织(图版Ⅰ-12)。4月上旬,造孢细胞停止分裂,转化为小孢子母细胞。由于细胞数目多,排列紧密,小孢子母细胞受挤压形成的棱角极为显著(图版Ⅰ-13)。同时,一些小孢子母细胞由于被挤压或吸收,逐渐退化并解体,这些退化的小孢子母细胞在小孢子囊内形成很大的空腔(图版Ⅰ-14)。小孢子母细胞进一步发育,彼此逐渐分离,此时,游离的母细胞呈多边形(图版Ⅰ-15)。
4月中旬,小孢子母细胞进入减数分裂时期。经过前期Ⅰ的细线期(图版Ⅰ-16)、偶线期(图版Ⅰ-17)、粗线期(图版Ⅰ-18)、双线期(图版Ⅰ-19)、终变期(图版Ⅰ-20)、中期Ⅰ(图版Ⅰ-21)、后期Ⅰ(图版Ⅰ-22)和末期Ⅰ(图版Ⅰ-23),产生2个子核,在2个子核之间不形成细胞壁,直接进行减数第2次分裂,减数分裂后形成的四分体有四面体形(图版Ⅰ-24,27)和左右对称形(图版Ⅰ-25,27),小孢子母细胞减数分裂过程中所发生的胞质分裂为同时型,即在减数分裂Ⅰ后不形成细胞壁,没有二分体时期。在第2次分裂完成后,在4个核之间产生壁,同时分隔为4个细胞(图版Ⅰ-26)。在同一个小孢子囊内,小孢子母细胞的发育表现出不同步现象,在减数分裂Ⅰ时,有的小孢子母细胞处于前期Ⅰ,有的则处于末期Ⅰ(图版Ⅰ-28)。4月下旬,四分体共同的胼胝质解体,释放出小孢子。
2.4 雄配子体的发育刚释放出的小孢子体积小,圆球形,核位于中央,其细胞壁较薄,中央有1个大细胞核,胞质浓厚,没有显著的液泡,为单核中央期(图版Ⅰ-29)。随着发育的进行,胞质内出现小液泡并不断融合形成大液泡,核逐渐被挤到边缘位置,靠近细胞壁,此时为单核靠边期(图版Ⅰ-30)。随后,小孢子进一步发育,染色较深,细胞质内积累大量的营养物质,细胞质外是一圈由小液泡不断融合形成的周围大液泡,细胞壁逐渐增厚,壁上有一个较薄而平滑的区域,为萌发孔(图版Ⅰ-31)。至此,形成了一个成熟的单核花粉粒,不具原叶细胞。
5月上中旬,杜松进入传粉季节,不同植株发育的不同步,花粉大量散放传粉大概持续2星期,集中在5月10日至15日。散粉后,花粉落在胚珠珠孔分泌的传粉滴上被带入胚珠中,落入胚珠珠心的花粉开始萌发,长出花粉管。胚性细胞也随之进入花粉管,花粉管萌发后不久,进入休眠阶段,此时的花粉管内仅有1个胚性细胞(图版Ⅰ-32,33)。
次年3月中下旬,萌发的花粉解除休眠,继续生长,进入珠心组织中。随着花粉管的伸长,胚性细胞于4月上中旬进行1次不均等的分裂,形成2个不等大的细胞:体积较大的是生殖细胞;体积较小的是管细胞。管细胞在花粉管的前端,生殖细胞紧随其后(图版Ⅰ-34,35)。4月下旬,花粉管继续深入珠心组织中,生殖细胞分裂形成1个不育细胞和1个精原细胞,不育细胞和管细胞大小相似,精原细胞较大(图版Ⅰ-36,37)。管细胞和不育细胞随着花粉管继续往珠心深处移动,精原细胞于5月中旬开始分裂形成2个精子,即2个雄配子(图版Ⅰ-38,39)。从花粉萌发到雄配子的形成大约需要12个月。
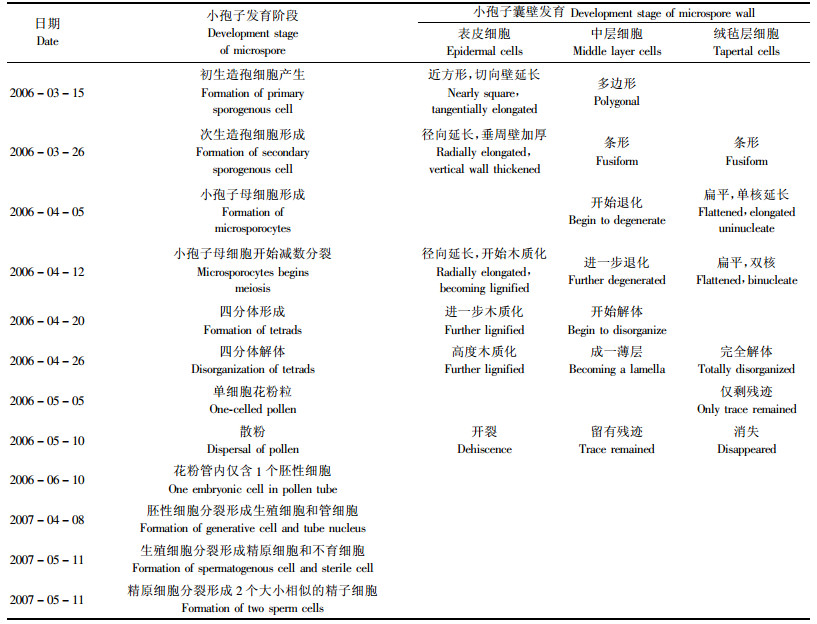
杜松小孢子发生及雄配子体发育进程见表 1。
|
|
在松柏类植物中,小孢子囊的起源有2种不同的发生方式:一种是孢子囊原始细胞由一群表面细胞组成,原始细胞平周分裂,形成小孢子囊壁、绒毡层和初生造孢细胞里面一层细胞;另一种是孢子囊原始细胞由一系列皮下细胞的平周分裂发生,小孢子叶的表皮包围在小孢子囊外(福斯特等,1983)。从小孢子囊的起源方式来看,杜松属于后一种发育类型。
在维管植物中,小孢子囊在小孢子叶上的着生位置有2种:一种着生在小孢子叶腋处;另一种着生在小孢子叶的下表面。在松柏目所有种中,小孢子囊是在小孢子叶下表面发育(福斯特等,1983)。而杜松小孢子囊在这2处位置都有发生。小孢子叶下表面着生2~4个小孢子囊;小孢子叶叶腋处的小孢子囊柄上着生1~8个小孢子囊,并且部分小孢子囊着生在更小的小孢子叶背面,杜松的小孢子叶球为复合小孢子叶球。这为裸子植物小孢子叶球的系统发育提供一个新的证据。
杜松传粉期的花粉为单核花粉粒,花粉内仅含1个胚性细胞。经传粉后,在胚珠珠心内经历1年的休眠时期,第2年春天,休眠后的花粉继续发育,胚性细胞在花粉管内才逐渐分裂形成管细胞、不育细胞和2个精子细胞。这种现象在秃杉(Taiwania cryptomerioides)和红豆杉科大部分植物中都有发生(王伏雄等,1980;陈祖铿等,1978)。
裸子植物小孢子母细胞一般经过冬季休眠,第2年春天开始分裂(陈祖铿等,1985;曹玉芳,1997)。而杜松的小孢子母细胞是在春天3月份开始形成造孢细胞,4月份形成小孢子母细胞,不经过休眠过程,就开始分裂。从造孢细胞开始形成,到小孢子散粉,仅2个多月的时间。穗花杉(Amentotaxus argotaenia)小孢子母细胞也不经过冬季休眠,在4月底才分化形成(陈祖铿等,1985)。这是曹玉芳等(1997)归纳的3种小孢子发育进程之外的又一种发育进程。
小孢子母细胞减数分裂时,二分体之间不形成细胞壁,直接进行第2次减数分裂,四分体的形成属于同时型。杜松的四分体有四面体形和左右对称形2种类型。
在小孢子母细胞形成以后,部分小孢子母细胞开始退化解体,在小孢子囊内形成空腔。太白红杉(Larix chinensis)和侧柏(Platycladus orientalis)等中也有这种现象(曹玉芳,1997;俞晓敏,2003)。这可能是孢母细胞的营养物质缺乏造成。在小孢子母细胞开始分裂之前,需要积累大量的营养物质,而小孢子囊内的营养物质有限,只有通过降解部分小孢子母细胞,来维持其他的小孢子母细胞继续发育。被降解的小孢子母细胞可能在形成过程中,由于其细胞内的营养物质相对较少,无法维持自身正常的发育被降解。
就小孢子母细胞发育过程来看,从减数分裂前期Ⅰ开始到四分体形成之前,不同细胞之间的发育水平在时间上存在明显的差异。而从四分体形成到花粉散出,发育基本上一致。这种现象的发生可能是因为减数分裂过程中需要的营养物质需要由绒毡层提供,而小孢子母细胞所接受的营养物质的先后顺序不同,从而产生个体的发育时差。从四分体形成到产生单核小孢子,小孢子细胞内拥有足够的营养物质,发育趋于同步,直到散粉时同时散出。由于坡位不同和海拔差异,不同植株间的发育在时间上也有所不同,在较早和较晚的植株间相差10 d左右。生长在低海拔、阳坡的植株发育较早,高海拔、半阳坡的植株发育较迟。
曹玉芳, 姚郭义, 计方, 等. 1997. 侧柏小孢子的发生和雄配子体的形成. 西北植物学报, 17(2): 163-169. DOI:10.3321/j.issn:1000-4025.1997.02.006 |
曹泽猷. 1993. 杜松种子萌发及育苗技术的研究. 林业科技通讯, 4: 6-9. |
陈祖铿, 王伏雄. 1978. 白豆杉的胚胎发育及其系统位置的商榷. 植物分类学报, 16(2): 1-10. |
陈祖铿, 王伏雄. 1985. 穗花杉的胚珠结构与雌、雄配子体的发育. 植物学报, 27(1): 19-25. DOI:10.3321/j.issn:1000-4025.1985.01.002 |
董希文, 赵启凯, 等. 2002. 杜松赤枯病发生规律及防治技术研究. 防护林科技, 2: 14-17. |
福斯特A. S., 小吉福德E. M. 著. 1983. 维管植物比较形态学. 李正理等译. 北京: 科学出版社.
|
李正理. 1996. 植物组织制片学. 北京: 北京大学出版社.
|
高艳平, 胡玉山. 2005. 干旱寒冷地区园林绿化优良树种杜松. 河北林业科技, 4: 108. |
孙殿宝, 李树森, 胡乙山. 2005. 杜松播种育苗技术. 防护林科技, 3: 114-115. DOI:10.3969/j.issn.1005-5215.2005.03.053 |
王伏雄, 李宪章, 陈祖铿. 1980. 秃杉的胚胎发育及其与杉科各属的比较. 植物分类学报, 18(2): 129-137. |
卫金圈. 2001. 杜松育苗技术. 山西林业, 4: 17-18. |
张剑斌, 戴凤海, 李志新, 等. 2000. 杜松赤枯病病原菌的研究. 防护林科技, 1: 26-30. DOI:10.3969/j.issn.1005-5215.2000.01.010 |
张国秀, 赵文革, 丁维力. 1994. 杜松球果营养成分研究初报. 中国林副特产, 29(2): 16-17. |
张少贞, 朱格麟, 郭守军, 等. 1996. 杜松挥发油化学成分与资源利用研究. 植物研究, 16(3): 363-367. |
俞晓敏, 赵桂仿. 2003. 太白红杉小孢子的发生和雄配子体的发育. 植物学通报, 20(5): 579-584. |
中国植物志. 2000. 北京: 科学出版社. 7: 379-380.
|
邹国林, 齐常明. 1996. 果实保湿浸种变温方法处理杜松种子. 北方园艺, 12: 65. |