 2009, Vol. 45
2009, Vol. 45文章信息
- 张国君, 李云, 何存成, 孙宇涵, 贺佳玉.
- Zhang Guojun, Li Yun, He Cuncheng, Sun Yühan, He Jiayu
- 四倍体刺槐不同叶龄叶片的营养及叶形变化
- Nutrition and Growth of Leaves at Different Leaf Ages in Tetraploid Robinia pseudoacacia
- 林业科学, 2009, 45(3): 61-67.
- Scientia Silvae Sinicae, 2009, 45(3): 61-67.
-
文章历史
- 收稿日期:2008-03-21
-
作者相关文章



2. 兰州市七里河区南北两山环境绿化工程指挥部 兰州 730050
2. Direction Administration of Environmental Afforestation of South-North Mountains, District of Qilihe Lanzhou 730050
四倍体刺槐(Tetraploid Robinia pseudoacacia)是北京林业大学于1997年从韩国引入我国的刺槐优良无性系。其具有可利用年限长、萌芽力强的优良特性,而且可以防止水土流失,改善生态环境;更重要的是其叶片宽大肥厚、蛋白质含量高、柔嫩多汁、适口性好(张国君等,2007)。
自将四倍体刺槐引入以来,刘涛等(2004)对其叶片粗蛋白和矿质营养进行了初步分析,肯定了其作为饲料树种的引种价值。北京林业大学四倍体刺槐课题组在分析四倍体刺槐叶粉的粗蛋白、钙、磷等常规成分含量及氨基酸组分的基础上,依据世界卫生组织和粮农组织(WHO/FAO)提出的氨基酸评分模式,利用必需氨基酸组分及必需氨基酸指数对其营养价值和饲用价值进行评定;同时采用模糊最优局势决策法对其营养价值进行综合评价;并对四倍体刺槐不同生长时期及不同部位的叶片营养成分和不同生长时期的氨基酸营养进行了分析研究(李云等,2006);进而结合四倍体刺槐营养和生物量的研究初步确定了其最佳采摘或刈割时期。但上述营养分析是以整个生长季节和整株叶片为基础的,是将叶片的叶龄、生长发育及物候期综合在一起考虑的。而Jérme等(2002)、Lin等(2007)、Oliveira等(1996)、Robert等(1996)已在其他植物中证明了叶龄、生长发育及物候期对叶片营养存在重要影响。目前,四倍体刺槐叶片的营养价值、叶层生物量、叶绿素含量和叶形生长发育与叶龄的关系以及他们相互之间的关系研究尚未见报道。因此,本文探索了四倍体刺槐在生长季不同叶龄叶片的营养价值、叶层生物量、叶绿素含量和叶形生长发育的变化规律以及他们相互之间的关系,从而为确定其最佳采摘或饲料林刈割周期提供更准确的理论依据,以期合理指导饲料生产。
1 材料与方法 1.1 试验地概况试验地位于延庆县风沙源育苗中心(40°27′N,115°50′E),土壤为沙壤土,属大陆性季风气候,属温带与中温带、半干旱与半湿润带的过渡带,年平均气温8.5 ℃,年降水量450~520 mm,光照充足,无霜期180 d左右。
1.2 试验材料试验林为2005年4月初定植的1年生四倍体刺槐苗,并于2006年3月初平茬后萌生的林分。在试验林内随机取60株于2006年7月7日再次平茬,并从7月22日开始在植株主干生长部位顶端处挂牌作标记(不同叶龄叶片的叶绿素含量是从7月12日开始挂牌标记),至9月25日植株生长末期止,期间每间隔10 d挂一次牌同时测量并采样,其中叶绿素和叶层生物量重复3次,叶片营养平行测定2次取平均值。具体调查和采样方法如下:将60棵试验株根据长势配对,其中30棵作为调查株,另外30棵作为采样株;并在30棵调查株的7月22日—8月2日时期叶层中上部标记2片叶,每次测量其叶长、叶宽和叶厚;同时每次在30棵采样株内随机选取3棵植株,分别采取本次叶龄全部叶片后混匀,并于9月25日分别单株(3棵)采取各个叶龄叶层(相邻两次挂牌标记日期内的全部叶片为一叶层)的全部叶片,立即称鲜质量,然后用四分法分别取单株各个叶龄叶层的50 g鲜叶,用于叶绿素的测定,将每个处理剩下的所有叶片分别干燥处理后用于营养的测定。
测定叶绿素、营养成分和生物量的叶片叶龄是叶层标记时间到采样时间9月25日之间的天数,具体时间段分别为:0~25 d叶龄叶片是8月31日—9月25日时期内的叶片;25~35 d叶龄叶片是8月22日—8月31日时期内的叶片;35~45 d叶龄叶片是8月12日—8月22日时期内的叶片;45~55 d叶龄叶片是8月2日—8月12日时期内的叶片;55~65 d叶龄叶片是7月22日—8月2日时期内的叶片。
不同形状的叶片叶龄是从标记时间7月22日开始算起,具体时间段分别为:0~10 d叶龄叶片是7月22日—8月2日时期内的叶片;10~20 d叶龄叶片是8月2日—8月12日时期内的叶片;20~30 d叶龄叶片是8月12日—8月22日时期内的叶片;30~40 d叶龄叶片是8月22日—8月31日时期内的叶片;40~55 d叶龄叶片是8月31日—9月15日内的叶片。
1.3 测定方法水分:直接干燥法(GB/T 6435-1986);粗蛋白质:凯氏法(GB/T 6432-1994);粗脂肪:索氏抽提法(GB/T 6433-1994);粗灰分:干灰化法(GB/T 6438-1992);酸性洗涤纤维(张丽英,2003):范氏酸性洗涤纤维(MAFIC-LAB/ METH 023-2001);中性洗涤纤维(张丽英,2003):范氏中性洗涤纤维(MAFIC-LAB/ METH 023-2001);钙:高锰酸钾滴定法(GB/T 6436-2002);总磷:分光光度法(GB/T 6437-2002);叶绿体色素(郝再彬等,2004):分光光度法。
1.4 数据分析用统计分析软件SPSS 13.0对数据进行分析。
2 结果与分析 2.1 叶片的营养含量变化 2.1.1 不同叶龄叶层的营养四倍体刺槐同一时期(9月25日)不同叶龄叶层的营养含量如表 1所示。由表 1可知:在同一时期内,随着叶龄的增长,叶片粗蛋白不断增高,在35~45 d叶龄时达到最大值,含量高达17.20%,随后随叶龄的增长叶片粗蛋白降低,在55~65 d叶龄时其含量降低为14.77%;叶片中性洗涤纤维和酸性洗涤纤维含量的变化趋势近乎一致,均从25~35 d叶龄开始升高,在45~55 d叶龄时达到高峰,然后随之降低。表 1测定结果也表明:叶片的粗脂肪、粗灰分和钙含量均随着叶龄的增长而增加,而磷含量在不同叶龄之间变化不大。其中,粗脂肪在55~65 d叶龄的含量为0~25 d叶龄的1.6倍;进一步计算不同叶龄叶片的钙磷比得出,其比值也随着叶龄的增长而增加,并在45~55 d叶龄时钙磷比(质量比)超过了7:1,即超出了动物饲料允许的范围。由于叶片的粗蛋白含量越高,纤维含量越低,钙磷比越低,其营养价值就越高。为此不难得出,当叶片在35~45 d叶龄范围内,其营养价值较好。
|
|

45~55 d和55~65 d 2个叶龄叶层的钙磷比均超过了动物饲料允许的范围7:1,与在同期取整株叶片分析的结果不同(李云等,2006),这是因为整株取样时各个叶龄叶层比例不同改变了营养成分的比值。另外,成熟叶片的粗脂肪较幼嫩叶片的含量高,与Bassey等(2001)对一种蕨类植物(Diplazium sammatii)叶片的研究结果相似。
2.1.2 不同生长时期相同叶龄叶片的营养不同生长时期10 d叶龄(采样时叶片的叶龄为10 d,如8月12日的叶片为8月2日—8月12日之间生长的所有叶片)叶片的营养含量见表 2。由试验结果可知:8月份各生长时期同叶龄叶片的各种营养成分含量均适合,其中粗蛋白含量均超过了23%,在8月2日甚至高达28.57%;钙磷比值均处于家畜日粮的最佳比值2:1范围内;粗脂肪各个时期均在3%~4%范围内波动,变化不大。在接近生长末期的9月15日,10 d叶龄叶片的粗蛋白含量较8月份减少很多,为18.34%;其钙磷含量也同时降低,但钙磷比反而升高到了2.5:1。可见,四倍体刺槐相同叶龄叶片的营养价值在生长中期优于生长末期。
|
|

不同叶龄叶层的营养分析结果表明:叶片在35~45 d叶龄范围内,其营养价值较好。然而饲料林的叶片采摘周期和刈割周期不仅要考虑其营养价值较好,同时要保证其生物量较大。四倍体刺槐不同叶龄叶层生物量的变化见图 1。由图中可以看出,叶层的鲜质量与干质量的变化趋势几乎一致,均从55~65 d叶龄(植株平茬再生后的第一叶层)缓慢增加,在35~45 d叶龄时叶层的鲜干质量达到最高峰,然后其急剧降低,在25~35 d叶龄时叶层的生物量降到了与55~65 d叶龄叶层的生物量几乎一样低的水平,随后随着叶龄的减小叶层生物量继续降低。

|
图 1 不同叶龄叶层的生物量变化 Figure 1 Variety of biomass of leaf layer at different leaf ages |
55~65 d叶龄叶层生物量低的原因可能是植株平茬后基部刚刚再生出来的叶片随着时间的推移,其上部的叶片越来越多,其本身正常的生长越来越受到限制,同时其年龄也越来越大,甚至老化;而25~35 d至0~10 d叶龄叶层生物量不高的缘故是叶片生长期短,生物量积累少。
2.3 叶片叶绿素含量的变化 2.3.1 不同叶龄叶层的叶绿素变化四倍体刺槐不同叶龄叶层的叶绿素含量见表 3。对叶绿素的试验结果进行方差分析得出,不同叶龄叶层的叶绿素a、叶绿素b和总叶绿素含量均在0.01水平上显著。同时,由表中数据变化规律可以看出,不同叶龄叶绿素a、叶绿素b和总叶绿素含量的变化趋势一致,均在25~35 d和55~65 d叶龄出现2个高峰,在65~75 d叶龄急剧下降。进一步进行多重差异比较可知,55~65 d叶龄叶片的叶绿素b和总叶绿素含量均极显著高于25~35 d叶龄叶片(P<0.01),叶绿素a含量显著高于25~35 d叶龄叶片(P<0.05);65~75 d叶龄叶片的叶绿素a含量最低,且与其他叶龄叶片差异极显著(P<0.01);其总叶绿素含量也低于其他叶龄叶片。另外,叶绿素a/b值被认为是一个良好的衰老生理指标,本试验测定的叶绿素a/b值随着叶龄的增加而逐渐下降,表明在叶片衰老过程中叶绿素a的破坏明显快于叶绿素b。
叶片的叶绿素含量在25~35 d叶龄和55~65 d叶龄出现了2个高峰值,在35~45 d叶龄和45~55 d叶龄叶片的叶绿素含量下降,这可能与叶片的物候期和环境温度有关;Hiroyuki等(2002)对极地柳(Polar willow)叶片的研究表明叶片的物候期和环境温度对叶片的光合作用有重大影响;Shirke(2001)对牧豆树(Prosopis juliflora)的研究也表明成熟叶片的光合能力高于幼嫩和衰老叶片,但具体原因还有待于进一步研究。
|
|

高等植物叶片所含叶绿素的数量不仅与叶片老嫩有关,而且与季节有关。四倍体刺槐不同生长时期10 d叶龄叶片的叶绿素含量见表 4。经方差分析表明:四倍体刺槐不同生长时期相同叶龄叶片的叶绿素a含量之间及总叶绿素含量之间存在显著差异(P<0.05),叶绿素b之间存在极显著差异(P<0.01)。进一步进行多重差异比较可得,10龄叶片在8月22日的叶绿素a、叶绿素b和总叶绿素含量均最低,且总叶绿素含量与其他生长时期之间存在显著差异(P<0.05);在8月12日叶片的叶绿素b和总叶绿素含量均达到了最高,而且显著高于其他生长时期(P<0.05),叶绿素a的含量也仅次于9月15日。
|
|

不同生长时期相同叶龄叶片的叶绿素含量在8月12日达到了最高,在8月22日却降到了最低,这也可能与温度有关。因为叶绿素的生物合成过程,绝大部分都有酶的参与。温度影响酶的活性,因而影响叶绿素的合成。一般来说,叶绿素形成的最适温度是30 ℃左右,最高温度是40 ℃。温度在8月初开始逐渐升高,叶片叶绿素的生物合成也相对增加,到8月12日左右时虽然高温开始抑制叶绿素的合成,但叶绿素的含量累加到了最大值,随后持续高温使叶绿素合成受抑,分解继续,所以8月22日叶绿素含量降到了最低点。
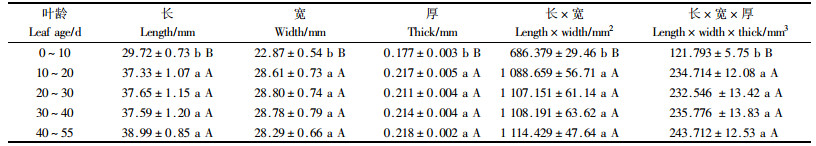
2.4 叶片形状的变化四倍体刺槐不同叶龄叶片的表观形状变化见表 5。由试验数据及方差分析可知,叶片的长、宽、厚、叶面积(长×宽)及叶形积(长×宽×厚)在叶龄之间均达到了极显著差异(P<0.01)。多重比较得出,四倍体刺槐0~10 d叶龄叶片的长、宽、厚、叶面积(长×宽)及叶形积(长×宽×厚)均极显著小于其他叶龄(P<0.01),而其他叶龄叶片的表观形状均无显著差异。这就表明叶片在10~20 d叶龄前生长迅速,10~20 d叶龄后表观形状生长缓慢。这也从侧面证明了为何叶片的营养含量、生长量和叶绿素含量的最大值出现在10~20 d叶龄以后。这为四倍体刺槐叶片的采摘周期和饲料林的刈割周期提供了理论依据。
|
|
四倍体叶片叶龄、营养含量、叶绿素含量及叶形之间的相关分析见表 6。结果表明:四倍体刺槐叶片叶龄与叶片粗脂肪、粗灰分和钙含量呈极显著正相关(P<0.01),与叶长和叶面积(长×宽)存在显著正相关关系(P<0.05);叶片叶绿素b和总叶绿素含量与叶片粗蛋白含量之间具有显著的负相关关系(P<0.05);叶长与叶片粗脂肪、粗灰分和钙含量呈显著正相关(P<0.05);叶面积(长×宽)与叶片粗脂肪和钙含量呈显著正相关(P<0.05),与叶片磷含量呈显著负相关(P<0.05);叶形积(长×宽×厚)与叶片粗灰分含量呈显著正相关(P<0.05)。
|
|

叶片粗脂肪和粗灰分的含量随叶龄的增大而增加,可能是叶片随叶龄的增大光合产物积累而增加的结果,这与谭忠奇等(2003)和Vijaya等(2006)对植物不同发育阶段叶片的研究结果相似。难以移动的元素Ca被林木地上部分吸收后,分配并被固定于植株叶部,所以随着叶龄越大叶片中钙含量越高。叶片随着叶龄的增大而生长发育,所以叶长和叶面积(长×宽)呈显著相关。正是由于叶片的生长发育和叶龄以及叶龄和叶片营养含量的上述关系,所以使得叶长与叶片粗脂肪、粗灰分和钙含量,叶面积(长×宽)与叶片粗脂肪和钙含量,叶形积(长×宽×厚)与叶片粗灰分含量均呈正相关;叶面积(长×宽)与叶片磷含量呈负相关。叶片叶绿素b和总叶绿素含量与叶片粗蛋白含量呈负相关,这与刘文彰等(1991)的研究结果相近;这可能是因为在生长季节的高温期叶绿素的生物合成受抑含量降低,而叶片蛋白含量由于光合产物的积累继续增加的结果。
2.5.2 不同生长时期相同叶龄叶片的营养、叶绿素含量之间的相关四倍体刺槐相同叶龄叶片的生育时期、营养含量及叶绿素含量之间的相关分析见表 7。从表 7可以看出,相同叶龄叶片的粗蛋白和磷含量与叶片生育时期之间存在显著的负相关关系(P<0.05);叶绿素b含量与叶片中性洗涤纤维和酸性洗涤纤维含量呈显著的正相关(P<0.05);总叶绿素含量与叶片中性洗涤纤维含量呈极显著的正相关(P<0.01)。
|
|

四倍体刺槐叶片粗蛋白含量与叶片生育时期呈显著负相关,即生长中期叶片粗蛋白含量高,生长末期由于粗蛋白发生转移而降低,这与先前的研究结果相同(李云等,2006;王改萍等,2006)。不同生育时期叶片磷含量的变化规律符合可移动矿质元素在阔叶树种中的一般变化规律(刘文彰等,1991),也与以前的研究结果相一致(黄宝龙等,1998;李云等,2006;王安友等,1996)。叶绿素与叶片纤维素含量呈显著相关的可能原因是,在一定范围内,叶绿素含量越多,光合越强,光合产物积累就越多,所以作为植物重要贮存物质的纤维素含量就越多。
3 结论与讨论1) 四倍体刺槐叶片的粗蛋白在同一时期内随叶龄的增大而增加,并在35~45 d叶龄达到最大值17.20%,然后迅速下降;中性洗涤纤维和酸性洗涤纤维含量变化趋势与粗蛋白近乎一致,只是最高峰出现在45~55 d叶龄;粗脂肪在55~65 d叶龄的含量为0~25 d叶龄的1.6倍;粗灰分和钙含量均随着叶龄的增长持续增加;综合叶片的各项营养成分含量得出,叶龄35~45 d以前的叶片营养价值较好。不同生长时期相同叶龄叶片的营养价值是生长中期好于生长末期。
2) 四倍体刺槐叶层生物量在35~45 d叶龄时达到最高峰,其次为45~55 d叶龄,而25~35 d叶龄与55~65 d叶龄叶层的生物量均较低。
3) 四倍体刺槐幼嫩叶片的叶绿素含量低,随着叶龄的增长,叶绿素含量不断增加,达到高峰,后来叶子衰老,叶绿素含量下降。不同生长时期相同叶龄叶片的叶绿素a、叶绿素b及总叶绿素含量之间也存在显著或极显著差异。
4) 四倍体刺槐不同叶龄叶片的长、宽、厚、叶面积(长×宽)及叶形积(长×宽×厚)在叶龄之间均达到了极显著差异,并且叶片在20 d叶龄前生长迅速,其后叶片表观形状生长缓慢。四倍体刺槐叶片叶龄与叶片粗脂肪、粗灰分和钙含量呈极显著正相关,与叶长和叶面积(长×宽)存在显著正相关关系;由于叶片的生长发育以及叶龄与叶片营养含量的上述关系,使得叶长与叶片粗脂肪、粗灰分和钙含量,叶面积(长×宽)与叶片粗脂肪、钙和磷含量,叶形积(长×宽×厚)与叶片粗灰分含量也存在相应的相关关系。
5) 四倍体刺槐相同叶龄叶片的粗蛋白和磷含量与叶片生育时期之间存在显著的负相关关系,叶绿素b含量与叶片中性洗涤纤维和酸性洗涤纤维含量呈显著的正相关,总叶绿素含量与叶片中性洗涤纤维含量呈极显著的正相关。
综合四倍体刺槐叶片营养价值随叶龄的变化趋势和生长中期高于生长末期的规律以及叶片的生长发育状况可以得出,当生长时期叶片在35~45 d叶龄内时,其营养价值较好,叶层生物量较高;这为四倍体刺槐叶片的采摘周期和饲料林的刈割周期提供了理论根据。四倍体刺槐不同叶龄的叶片叶绿素含量在35 d和65 d叶龄2个峰值之间有所下降,相同叶龄叶片的叶绿素含量在8月22日最低,而8月12日却达到了最高,可能与叶片的生育期和叶绿素的合成因素有关,其中主要是温度对叶绿素的影响,但具体原因还有待于进一步研究。另外,分析结果还表明:四倍体刺槐不同叶龄叶片的叶绿素b和总叶绿素含量与叶片粗蛋白含量呈负相关。这可能也与高温期叶绿素的生物合成受抑而使其含量降低的原因有关,也需以后深入研究。
郝再彬, 苍晶, 徐仲, 等. 2004. 植物生理实验. 哈尔滨: 哈尔滨工业大学出版社, 46-49.
|
黄宝龙, 叶功富, 张水松, 等. 1998. 木麻黄人工林营养元素的动态特性. 南京林业大学学报, 22(2): 1-4. |
李云, 张国君, 路超, 等. 2006. 四倍体刺槐不同生长时期和部位的叶片的饲料营养价值分析. 林业科学研究, 19(5): 580-584. DOI:10.3321/j.issn:1001-1498.2006.05.007 |
刘涛, 李春燕, 王莉. 2004. 西藏引种四倍体刺槐与普通刺槐营养成分对比分析. 中国野生植物资源, 23(2): 46. DOI:10.3969/j.issn.1006-9690.2004.02.017 |
刘文彰, 王双贵. 1991. 生长旺季刺槐叶片营养成分含量及其变化研究初报. 河北师范大学学报, (4): 43-46. |
谭忠奇, 林益明, 向平, 等. 2003. 5种榕属植物不同发育阶段叶片的热值与灰分含量动态. 浙江林学院学报, 20(3): 264-267. DOI:10.3969/j.issn.2095-0756.2003.03.009 |
王安友, 任莉. 1996. 银杏叶营养元素季节性变化规律的研究. 林业科学研究, 9(2): 133-137. |
王改萍, 彭方仁, 李生平. 2006. 银杏叶片蛋白质含量动态变化的电泳分析. 南京林业大学学报, 30(4): 114-118. DOI:10.3969/j.issn.1000-2006.2006.04.027 |
张国君, 李云, 姜金仲, 等. 2007. 饲料型四倍体刺槐叶粉饲用价值的比较研究. 草业科学, 24(1): 26-30. DOI:10.3969/j.issn.1001-0629.2007.01.006 |
张丽英. 2003. 饲料分析及饲料质量检测技术. 第2版. 北京: 中国农业大学出版社, 63-70.
|
Bassey M E, Etuk E U I, Ibe M M, et al. 2001. Diplazium sammatii: Athyriaceae ('Nyama Idim'): Age-related nutritional and antinutritional analysis. Plant Foods for Human Nutrition, 56: 7-12. DOI:10.1023/A:1008185513685 |
Hiroyuki M, Masaki U, Masako M, et al. 2002. Leaf photosynthetic characteristics and net primary production of the polar willow (Salix polaris) in a high arctic polar semi-desert, Ny-Alesund, Svalbard. Canadian Journal of Botany, 80: 1193-1202. DOI:10.1139/b02-108 |
Jérme O, Jean-Marc O, Gilles B. 2002. Seasonal and spatial patterns of foliar nutrients in cork oak (Quercus suber L.) growing on siliceous soils in Provence (France). Plant Ecology, 164: 201-211. |
Lin Y M, Liu J W, Xiang P, et al. 2007. Tannins and nitrogen dynamics in mangrove leaves at different age and decay stages (Jiulong River Estuary, China). Hydrobiologia, 583: 285-295. DOI:10.1007/s10750-006-0568-3 |
Oliveira G, Martins-Louçâo M, Correia O A, et al. 1996. Nutrient dynamics in crown tissues of cork oak (Quercus suber L.). Trees, 10: 247-254. |
Robert B, Caritat A, Bertoni G, et al. 1996. Nutrient content and seasonal fluctuations in the leaf component of cork oak (Quercus suber L.) litterfall. Vegetatio, 122: 29-35. DOI:10.1007/BF00052813 |
Shirke P A. 2001. Leaf photosynthesis, dark respiration and fluorescene as influenced by leaf age in an evergreen tree, Prosopis juliflora. Photosynthetica, 39(2): 305-311. DOI:10.1023/A:1013761410734 |
Vijaya S, Charles K P, Dhananjay S. 2006. Phosphorus nutrition and tolerance of cotton to water stress:Ⅰ. Seed cotton yield and leaf morphology. Field Crops Research, 96(2/3): 191-198. |