 2009, Vol. 45
2009, Vol. 45文章信息
- 张莉燕, 盛建东, 武红旗, 朱建雯, 高军.
- Zhang Liyan, Sheng Jiandong, Wu Hongqi, Zhu Jianwen, Gao Jun
- 新疆柽柳立地土壤养分的空间变异特征
- Soil Nutrient Characteristic of Tamarix site in Xinjiang
- 林业科学, 2009, 45(3): 54-60.
- Scientia Silvae Sinicae, 2009, 45(3): 54-60.
-
文章历史
- 收稿日期:2008-04-01
-
作者相关文章



土壤是变化的,包括土壤随时间和空间位置发生的变化,土壤随空间位置发生的变化,称作土壤的空间变异(秦耀东,2003)。“肥岛”是干旱、半干旱区土壤养分在生物、非生物过程作用下富集于灌木周围,使土壤养分由冠幅中心向冠幅外逐渐递减的现象。灌木“肥岛"是荒漠生态系统中的一种普遍现象,也是荒漠生态系统中独特的地理单元——生物地球化学循环最活跃的区域。灌木“肥岛"是植物与多种环境因子交互作用的结果,其形成与发育又对荒漠植被具有重要的反馈作用,因此干旱区灌木“肥岛"现象从群落生态学到全球变化生态学等领域都受到了广泛的关注。从研究区域来看,目前有关灌木“肥岛"的研究主要集中在北美(Hook et al., 1991);从研究方法来看,大多研究集中在“肥岛"与“岛"外土壤要素的对比。在国内,陈广生等(2003)、张宏等(2001)、吕贻忠等(2003)从不同角度分别就干旱和半干旱区灌木下“肥岛"研究方法、意义和干旱草地灌丛化与土壤异质性关系方面的研究进行了总结。
新疆地处内陆干旱区,独特的生物气候和封闭的地形,发育了独具特色的荒漠植被。新疆是我国柽柳属(Tamarix)植物种类最多(16种)、分布最广的省区(约533万hm2),占全国柽柳种类的80%。柽柳属显域灌丛,在南北疆均有大面积分布。柽柳属植物具抗旱、耐盐碱、耐水湿、耐沙埋及耐贫瘠的特性,在干旱、半干旱地区始终保持着其优势种和建群种的地位(张道远等,2003;陈晓琴等,2006)。这些植被在防风固沙、维护生态安全、改善生态环境方面发挥着巨大的作用。在荒漠植被与环境条件的相互作用下,形成土壤的空间变异性,由于该区域属于世界典型的干旱与极端干旱区,植被以旱生和超旱生为主,该区域的“肥岛”现象的研究还是空白。
为此,本文利用地统计学和GIS技术,对荒漠植被的“肥岛"特征、作用范围,植被消亡与环境变化的关系进行研究,旨在阐明“肥岛”在荒漠生态系统中的形成机制和生态学意义,为动态监测干旱区荒漠环境的变化提供理论依据与基础资料,也为灌木-荒漠生态系统的结构和功能的演替和在环境变化中的作用提供一些参考依据。
1 研究地区概况与研究方法 1.1 研究区概况 1.1.1 南疆塔里木河自然概况该研究区选在轮台县的轮南镇,轮南镇地处新疆塔里木河(简称塔河)的中游,塔克拉玛干大沙漠北缘,属典型的暖温带大陆性荒漠气候。该区植被主要以灌木和半灌木群落为主,除了短命植物和1年生植物外,几乎全为旱生和超旱生植物,具有典型荒漠植被的特点。群落以柽柳和胡杨(Populus euphraticu)为主。研究区的地理位置41.275°—41.280° N,84.207°—84.213° E,海拔918~920 m。全年平均降水量41 mm,年均蒸发量2 778 mm,4—7月多大风,并伴有风沙浮尘,主风向为北偏西,年平均风速2.3 m·s-1,最大风速为30 m·s-1。
1.1.2 北疆阜康自然概况该研究区选在阜康市的阜北农场,属大陆性中温带气候,位于天山北麓,准噶尔盆地南缘,南依天池风景区,北接古尔班通古特沙漠,地势开阔平坦,四季分明,以旱生的灌木、半灌木以及肉质植物为主,泌盐、储水、高渗透压、枝叶肉质化、枝叶极度缩小等植物的生态适应现象普遍存在,具有典型荒漠植被的特点。研究区的地理位置44.269°—44.271° N,88.048°—88.049° E,海拔479~485 m。年均降水量150 mm,年蒸发潜力在2 000 mm以上,山区年均风速为2.9 m·s-1,平原、荒漠区分别为2.4和2.6 m·s-1。
1.2 研究方法 1.2.1 样品采集2006年10月3号,在塔河中游选取人为干扰小的典型样区进行采取,分别进行低密度、高密度采样。低密度样是选取冠幅、株高和生长发育情况基本一致的灌丛,分别按大、中、小和枯死的4个生长阶段分类采样(表 1),大的取了8丛,小的和中的各取9丛,枯死的取了5丛。每丛以冠幅中心(根部)、冠幅边缘(树冠垂直投影边缘)、冠幅外(树冠垂直投影外)的同心圆采样法,在表层0~20 cm深度的4个方向各采1个混合土样。高密度样是对一丛大柽柳,从冠幅中心向正东、正南、正北、正西4个方向,除了中心点,各个方向各取表层0~20 cm深度的12个点,前7个点以50 cm为间距,后5个点以1 m为取样间距,共计49个土样。植株位置采用GPS定位。
|
|
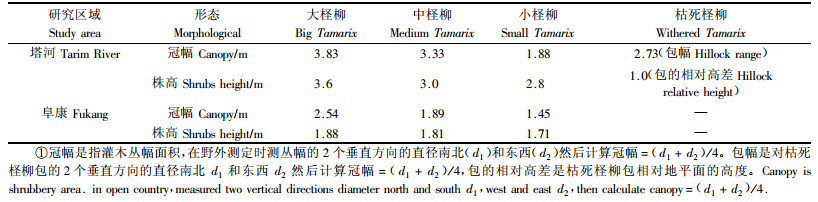
2007年5月3号,在阜康市的阜北农场也采取了低密度、高密度样。低密度样采取了大、中、小3个生长阶期作对比,各取了5丛。高密度样也在4个方向,除了中心点,各个方向各取12个点,前8个点以30 cm为间距,后4个点以50 cm为取样间距,即49个土样(表 1)。
1.2.2 土壤分析方法土壤样品经自然风干,过筛后进行室内分析,分析方法采用常规方法。有效磷的测定:0.5 mol·L-1 NaHCO3浸提,钼锑抗比色法。速效钾的测定:1.0 mol·L-1 NH4OAc浸提,火焰光度法。水解氮的测定:1.8 mol·L-1 NaOH碱解扩散法。有机质的测定:重铬酸钾容量法(外加热法)。
1.2.3 数据处理利用富集率(enrichment ratio,E)来表征土壤养分富集程度。EA=A/C,反应柽柳根部“肥岛”效应;EB=B/C,反应柽柳冠幅边缘养分的富集程度,其中A表示冠幅中心,B表示冠幅边缘,C表示冠幅外。利用SPSS 11.5软件进行统计分析,采用LSD法对同一发育阶段不同采样部位土壤养分的差异进行多重比较;采用单因素方差分析法(one-way ANOVA)比较不同发育阶段同一取样部位的富集率的差异。高密度样用GS+ for windows 5.0进行半方差分析,采用Surfur 7.0地统计软件制图并进行Kriging空间插值。
2 结果与分析 2.1 柽柳“肥岛”的土壤养分特征及富集率经鉴定塔河柽柳是多枝柽柳(Tamarix ramosissina),阜康柽柳是短穗柽柳(Tamarix java)。由表 2有机质含量可以看出,2个研究区柽柳冠幅中心有机质含量与冠幅边缘和冠幅外呈极显著差异,而冠幅边缘与冠幅外差异不显著。塔河水解氮冠幅中心与冠幅边缘差异极显著而与冠幅外不显著,冠幅边缘与冠幅外差异也不显著,阜康水解氮冠幅中心极显著高于冠幅边缘和冠幅外。塔河柽柳下的有效磷冠幅中心与冠幅外差异显著,与冠幅边缘差异不显著。阜康只有大柽柳冠幅中心与冠幅边缘和冠幅外差异极显著,其余都不显著。塔河大柽柳的速效钾冠幅中心极显著大于冠幅边缘和冠幅外,冠幅边缘与冠幅外差异不显著,中柽柳和小柽柳速效钾冠幅中心与冠幅边缘和冠幅外差异显著。阜康大柽柳冠幅中心和冠幅边缘与冠幅外差异极显著,中柽柳和小柽柳冠幅中心、冠幅边缘和冠幅外的差异均达极显著水平。
|
|
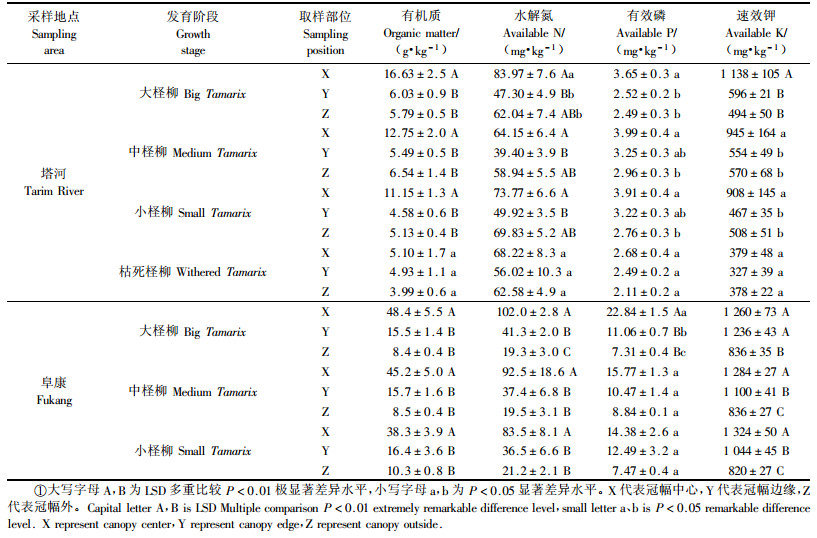
从整体均值看出,2个研究区柽柳冠幅中心土壤养分含量与冠幅边缘或冠幅外差异显著,而冠幅边缘与冠幅外差异大都不显著,但塔河枯死柽柳的土壤养分都呈不显著性。阜康柽柳立地土壤养分普遍高于塔河。塔河土壤养分含量有的是冠幅中心>冠幅边缘>冠幅外,有的是冠幅中心>冠副外>冠幅边缘。而阜康土壤养分含量均为冠幅中心>冠幅边缘>冠幅外,呈现出规律性的递减趋势,显示出较明显的“肥岛”效应。
由表 3看出,柽柳根部的富集率高于冠幅边缘(EA和EB>1则表示富集),其中塔河柽柳根部土壤养分的富集率都大于1,冠幅边缘富集率有的大于1,有的小于1,而阜康柽柳根部和冠幅边缘土壤养分的富集率都大于1,并且有机质、水解氮和有效磷普遍比塔河的富集率高。这可能与2个研究区的地貌特征和气候差异较大有关,塔河柽柳呈包状生物地貌,这种地貌特征主要是由于该区域风沙较大,生物富集与风蚀沙化并存,物质在富集的同时又被沙粒覆盖,这样使得“肥岛”在水平方向上的富集强度减弱,而在垂直方向延伸,加之柽柳包坡度的影响,养分资源不易在冠幅下累积。而阜康柽柳地表平坦且风沙相对较小,容易使土壤养分由冠幅外向冠幅内聚集。
|
|
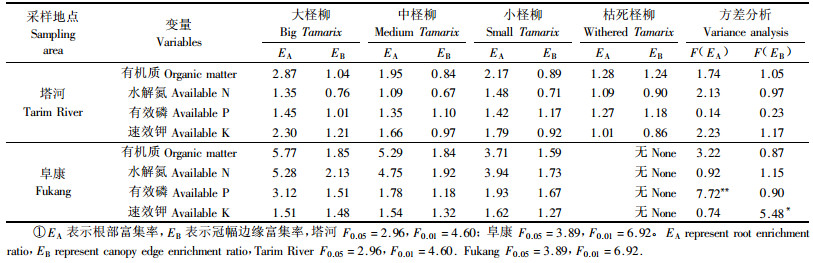
2个地区不同发育阶段的柽柳的养分富集率除了有效磷大柽柳有极显著差异外,其他各养分基本没有达到显著差异。这表明柽柳的植株发育阶段对“肥岛”的强度影响较弱。
2.2 柽柳“肥岛”的有效作用范围由表 4可以看出塔河和阜康土壤养分的变异系数在21%~82%之间,为中等变异(CV<10%为弱变异性,CV=10%~100%为中等变异性,CV>100%为强变异性(王政权,1999)且符合正态分布。
|
|
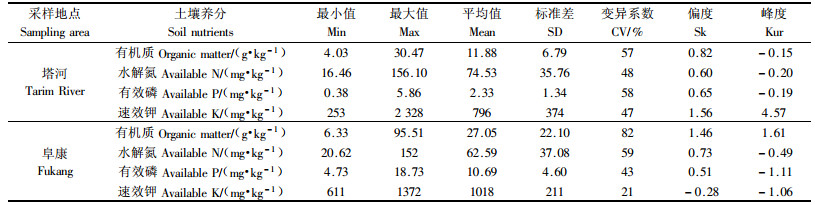
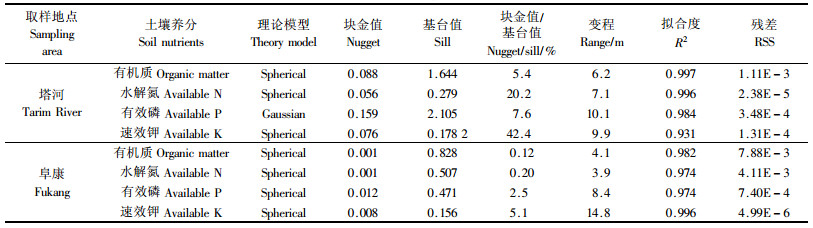
由表 5看出,除塔河有效磷符合高斯模型外,其余都符合球状模型,R2达到极显著水平,且残差也很小,说明拟合模型有效。塔河有机质的变程是6.2 m,有效磷是10.1 m,速效钾是9.9 m,水解氮是7.1 m,冠幅是3.82 m。阜康土壤养分的变程有机质是4.1 m,有效磷是8.4 m,速效钾是14.8 m,水解氮是3.9 m,冠幅是2.61 m。在各自的变程范围内,土壤属性值之间存在着一定的相关关系,如大于该值时,则说明它们之间是相互独立的。除塔河的速效钾块金值与基台值的比值大于25%外,两地其余土壤养分的块金值与基台值的比值在0.12%~20.0%之间,说明土壤养分具有强烈的空间相关性,且其变异主要是由结构性因素引起的(当比值小于25%,说明系统具有强烈的空间相关性;如果比例在25%~75%之间,表明系统具有中等的空间相关性;大于75%说明系统空间相关性很弱(王政权,1999),这与采样区为自然荒漠区,受人为干扰弱,并且与地形、质地、降水等因素一致,所以此变异主要是荒漠植被的“肥岛”作用引起的。
|
|
利用地统计学方法对各养分含量进行克里格插值(图 1,2,3,4),可以看出土壤养分由根部向周围呈递减趋势,表现出相对明显的“肥岛”效应。塔河的有效磷向东南向偏高,速效钾向北偏高。阜康有效磷向西升高,速效钾朝西南向升高。这可能与当地的风向、地形或地表径流有关。

|
图 1 土壤有机质克立格插值图 Figure 1 Kriging interpolation map of the soil organic matter (0,0)点为根部中心。横坐标和纵坐标的单位:m。下同。 (0, 0)spot is the root center. Abscissa and y-coordinate unit:m. The same below. |

|
图 2 土壤水解氮克立格插值图 Figure 2 Kriging interpolation map of the soil available N |

|
图 3 土壤有效磷克立格插值图 Figure 3 Kriging interpolation map of the soil available P |

|
图 4 土壤速效钾克立格插值图 Figure 4 Kriging interpolation map of the soil available K |
1) 阜康柽柳的冠幅和株高极显著小于塔河的柽柳,土壤养分却远远高于塔河。虽然2个地区的柽柳为不同的种,但这主要与南北疆气候条件有关,年均降水量北疆阜康(150 mm)大于南疆塔河(41 mm),生物富集作用强。同时南疆的风沙吹蚀和填埋作用,形成柽柳包,降低了养分的富集强度。
2) 在干旱、半干旱的荒漠生境下灌丛柽柳为了适应极端恶劣的生存环境,有效地利用养分,形成了“肥岛”效应(Garner et al., 1998;Schlesinger,1996;Schlesinger et al., 1990)。柽柳立地土壤养分冠幅中心高于冠幅边缘和冠幅外,而冠幅边缘与冠幅外大都没有明显差异,这与李君等(2007)、张强等(2006)研究结果一致。也可看出不同灌木对养分聚集的强度不同,且同一灌木在不同的生境下也存在着差别。而塔河枯死柽柳的土壤养分差异不显著,表明灌木死亡后“肥岛”效应逐渐消退,同时也说明冠幅、根系、枯落物及其微生物等生物因素在灌木“肥岛”形成中的重要作用,一旦植株死亡,所富积的有限养分资源很快就矿化分解或被风蚀沙化。因此,保护荒漠生态系统的植被资源对于减少养分资源尤其是有机碳的释放具有重要作用。
3) 塔河柽柳“肥岛”的作用范围在6.2~10.1 m的范围。阜康的在4.1~14.8 m的范围内。阜康取样在4.4 m的范围内,塔河取样在8.5 m的范围内,而阜康柽柳冠幅是2.61 m,塔河柽柳冠幅是3.82 m。这说明柽柳的有效作用范围在冠幅之外,且取样尺度比较合理(盛建东等,2003)。塔河和阜康的块金值与基台值的比值除塔河的速效钾变异系数大于25%外,两地其余土壤养分的块金值与基台值的比值小于25%,说明具有强烈的空间相关性,且其变异主要是由结构性因素引起的。这与采样区为自然荒漠景观,受人为干扰弱,并且与地形、质地、降水等因素相对一致,所以此变异主要是“肥岛”作用引起的。
4 结论1) 在两研究区中,柽柳立地土壤养分冠幅中心高于冠幅边缘和冠幅外,而冠幅边缘与冠幅外没有明显差异。塔河枯死柽柳的土壤养分差异不显著。
2) 柽柳土壤养分的富集率都大于1,养分是富集的。塔河枯死柽柳的土壤养分富集率也大于1,表明“肥岛”的形成和消逝都是长期过程。2个地区不同发育阶段的柽柳的养分富集率没有达到显著差异。
3) 塔河柽柳“肥岛”的作用范围在6.2~10.1 m之间,阜康柽柳“肥岛”的有效作用范围在4.1~14.8 m之间,且在冠幅之外。研究区柽柳立地土壤养分具有强烈的空间相关性。
研究结果对于了解新疆柽柳“肥岛”特征以及揭示荒漠植物“肥岛”形成机制提供可靠的数据支撑,同时对植物-土壤相互作用提供理论依据。
陈广生, 曾慧德, 陈伏生, 等. 2003. 干旱和半干旱地区灌木下土壤"肥岛"研究进展. 应用生态学报, 14(12): 2156-2161. |
陈晓琴, 王婷, 汪建红. 2006. 新疆柽柳属植物的价值及开发利用建议. 新疆师范大学学报:自然科学版, 25(3): 100-102. |
李君, 赵传义, 朱宏. 2007. 梭梭和柽柳的"肥岛"效应. 应用生态学报, 17(12): 5137-5138. |
吕贻忠, 马兴旺. 2003. 荒漠化土壤养分变化的影响因素研究进展. 生态环境, 12(4): 473-477. DOI:10.3969/j.issn.1674-5906.2003.04.025 |
秦耀东. 2003. 土壤物理学. 北京: 高等教育出版社.
|
盛建东, 杨玉玲, 陈冰, 等. 2003. 克拉玛依干旱生态农业区土壤有机质与全氮空间变异特征研究. 新疆农业大学学报, 26(4): 32-36. DOI:10.3969/j.issn.1007-8614.2003.04.008 |
王政权. 1999. 地统计学在生态学中的应用. 北京: 科学出版社, 15-30.
|
张宏, 史培军, 郑秋红. 2001. 半干旱地区天然草地灌丛化与土壤异质性关系研究进展. 植物生态学报, 25(3): 366-370. DOI:10.3321/j.issn:1005-264X.2001.03.017 |
张强, 程滨, 杨治平, 等. 2006. 芦芽山鬼箭锦鸡儿灌丛营养特征及土壤养分分布规律. 应用生态学报, 17(12): 229-231. |
张道远, 尹林克, 潘伯荣. 2003. 柽柳属植物抗旱性能研究及其应用潜力评价. 中国沙漠, 23(3): 252-256. DOI:10.3321/j.issn:1000-694X.2003.03.008 |
Garner W, Steinberger Y. 1998. Approposed mechanism for the formation of 'fertile islands' in the desert ecosystem. J Arid Environ, 16: 257-262. |
Hook P B, Burke I C, Lauenroth W K. 1991. Heterogeneity of soil and plant N and C associated with individual plants and opening in North American shortgrass steppe. Plant Soil, 138: 247-256. DOI:10.1007/BF00012252 |
Schlesinger W H. 1996. On the spatial pattern of soil nutrients in desert ecosystem. Ecology, 77(2): 364-374. |
Schlesinger W H, Reynolds J F, Cunningharm G L, et al. 1990. Biological feedbacks in global desertification science. Ecology, 247: 1043-1048. |