 2009, Vol. 45
2009, Vol. 45文章信息
- 姜英淑, 陈书明, 王秋玉, 卢宝明.
- Jiang Yingshu, Chen Shuming, Wang Qiuyu, Lu Baoming
- 碳酸盐和干旱胁迫对欧李生理特性的影响
- Effect of NaHCO3 and Drought Stress on Physiological Characteristics of Cerasus humilises
- 林业科学, 2009, 45(3): 19-23.
- Scientia Silvae Sinicae, 2009, 45(3): 19-23.
-
文章历史
- 收稿日期:2008-08-28
-
作者相关文章



2. 北京市林业种子苗木管理总站 北京 100209;
3. 三门峡职业技术学院 三门峡 472000
2. General Administration of Forest Seeds and Seedlings of Beijing Beijing 100029;
3. Sanmenxia Professional Technology Institute Sanmenxia 472000
欧李(Cerasus humilis )是我国特有的一种蔷薇科(Rosaceae)樱桃属(Cerasus)矮小灌木果树,根系发达,耐旱、抗寒、耐贫,并有大量落叶归还于土壤,是不可多得的绿化荒山、改良土壤、荒漠化治理及防止水土流失的树种。20世纪80年代以来,我国科研工作者对欧李进行了大量研究,包括种质资源调查与收集(王有信等,2005)、生物学特性(奥小平,2006)、杂交育种与品种培育(王有信等,2007)、优良品种繁殖与栽培技术(姚岘武等,2007;庄丽娟等,2005)、果实品质与加工(马建军等,2008;李志国等,2005)等。但对欧李抗逆性研究的报道很少(张毅等,2003;楮建平等,2008)。
本研究对欧李进行高浓度的碳酸盐处理、干旱处理以及盐与干旱共同处理,探讨其在碱盐和干旱条件下的多个理化指标的变化特征,掌握欧李耐盐碱性和耐旱性的理化机制,为欧李品种选育奠定基础。
1 材料与方法 1.1 试材及处理2006年初将2年生黄山店欧李实生苗(由北京市林业种子苗木管理总站提供)移栽到东北林业大学林木育种基地塑料大棚内,定植于直径30 cm、高25 cm的花盆中,每盆1株。试验于2007年7—8月在大棚内进行,材料为3年生实生苗。共设置4种处理:1)对照给苗木浇清水至饱和,以后每天持续正常浇清水;2)盐处理给苗木浇清水至饱和,以后每隔2 d浇1次300 mmol·L-1的NaHCO3溶液300 mL,同时每天浇适量清水;3)干旱处理给苗木浇清水至饱和,以后每隔2 d浇灌清水300 mL;4)盐与干旱共同处理给苗木浇清水至饱和,然后每隔2 d浇1次300 mmol·L-1的NaHCO3溶液300 mL。每个处理4棵植株,3次重复,共处理25 d,第26天开始复水解除胁迫,即正常浇水至饱和。分别在处理过程中的第3,6,9,12,15,20,25和31天对试材分处理和单株取叶片样品进行理化指标测试,第31在数据为复水后数据。
1.2 测定方法测定的指标有电解质外渗率、叶绿素a含量、叶绿素a含量与叶绿素b含量比值、类胡萝卜素含量、类胡萝卜素与总叶绿素含量比值、丙二醛(MDA)含量、超氧化物歧化酶(SOD)和过氧化物酶(POD)活性和脯氨酸含量。电解质外渗率、SOD、POD和MDA的测定参照郝再彬等(2004)的方法;用95%乙醇浸提法提取叶绿素与类胡萝卜素;茚三酮比色法(张志良等,2004)测定脯氨酸含量。
1.3 数据处理对测得数据的平均值进行方差分析和相关分析,百分数据进行反正弦转换。采用Microsoft Excel软件作图。
2 结果与分析 2.1 电解质外渗率和MDA含量盐、干旱胁迫对欧李电解质外渗率的影响如图 1。盐处理植株在整个处理期间电解质外渗率没有明显变化且低于对照,在前9天,干旱处理和盐与干旱共同处理植株的电解质外渗率变化平缓且略低于对照,第9~25天呈先升高后降低趋势,复水后,各处理的电解质外渗率与对照相等。由试验结果不难看出,单一的盐处理对欧李的细胞膜透性没有造成明显的伤害;在干旱和盐与干旱共同胁迫条件下,细胞膜渗透率随处理时间延长呈一个从相对稳定期向不断升高后下降的趋势,细胞膜完整性由前期的相对完整向后期的破损坏死转变,细胞受损伤的程度在处理后期开始加剧。

|
图 1 逆境胁迫对电解质外渗率与丙二醛含量的影响 Figure 1 Effect of the stress on electrolyte exosmosis percentage and MDA content  对照Controol; 对照Controol; 盐处理NaHCo3 stress; 盐处理NaHCo3 stress; 盐与干旱期处理NaHCO3 and drought stress; 盐与干旱期处理NaHCO3 and drought stress; 干旱处理Drought stress.下同The same below. 干旱处理Drought stress.下同The same below. |
植物的细胞膜主要由磷脂和蛋白质构成,磷脂在活性氧存在的情况下易被过氧化,进而改变膜结构、破坏膜原有的生物学功能。丙二醛(MDA)是膜脂过氧化的重要产物,MDA从膜上释放后,可以与蛋白质、核酸反应,改变这些大分子的构型,或使之产生交联反应,从而丧失功能,还可使纤维素分子间的桥键松驰,或抑制蛋白质的合成。因此,MDA的积累可能对膜和细胞造成一定的伤害,所以含量的变化可以反映细胞膜受损程度。图 1还显示,欧李MDA含量在整个处理期间表现为:对照处理波动不大;盐与干旱共同胁迫和单一干旱胁迫在处理的前6天大幅下降,单一碳酸盐胁迫处理略有上升,第6~20天各处理均先升高后降低,第25天及复水后变化比较平缓且均低于对照;盐处理植株的MDA含量在处理中期高于对照、干旱处理和盐与干旱协同处理,这也说明欧李的细胞膜对盐处理更敏感;而胁迫后期,盐处理细胞膜的电解质外渗率和MDA含量低于并接近对照,而干旱处理和盐与干旱共同处理均明显低于对照,这说明欧李可能具有较强的碳酸盐适应性。
2.2 保护酶活性图 2显示,盐处理下欧李的POD活性在整个处理期间变化比较平缓并且均略高于对照;盐与干旱协同处理和干旱处理呈先升高后降低趋势,干旱处理的降低趋势大于盐与干旱协同处理,且第12天以后均低于对照。说明单一的碳酸盐胁迫、轻度的干旱胁迫和盐与干旱协同处理都能激发POD活性,但是随着胁迫时间的延长,活性氧及其产物抑制了保护酶的合成、加速了酶的分解,从而导致POD活性下降,特别是干旱处理和盐与干旱共同处理。

|
图 2 逆境胁迫对POD与SOD活性的影响 Figure 2 Effect of the stress on POD and SOD activity |
图 2还显示,各个处理的SOD活性在整个处理期间变化平缓,各处理间SOD活性没有明显差异,复水后各处理SOD活性均呈上升趋势,且均高于对照,盐与干旱协同胁迫的上升幅度最大。也就是说在不同胁迫处理期间,SOD不是主要的体内活性氧清除物质,而胁迫解除后,SOD在清除植物体内活性氧方面起到了主要作用。
2.3 叶绿素和类胡萝卜素叶绿素含量是反映植物光合强度的重要指标。图 3显示,在处理的前10天,各处理叶绿素a变化平缓,第12~15天各处理均急剧上升,但处理间差异不大,第15~25天各处理间存在明显差异,且均低于对照,复水后呈上升趋势,干旱胁迫上升的幅度最大,说明由渗透胁迫对叶绿素a造成的伤害可能容易修复,由离子造成的伤害修复缓慢。

|
图 3 逆境胁迫对叶绿素a及叶绿素a含量与叶绿素b含量比率的影响 Figure 3 Effect of the stress on chlorophyll a content and ratio between chlorophyll a and chlorophyll b content |
叶绿素a含量与叶绿素b含量比值可以反映叶绿体中类囊体的跺叠程度,即不同堆叠状态的类囊体比例。叶绿素a含量与叶绿素b含量比值越低,类囊体的堆叠程度越低。类囊体膜的稳定性也越差,致使光能在2个光系统之间的分配受到影响,从而降低光合作用(赵文恩等,1999)。图 3还显示,胁迫期间各处理的叶绿素a含量与叶绿素b含量比值呈缓慢下降趋势,其中盐与干旱协同处理的变化幅度较大,处理的第20~25天该值明显低于对照和盐或干旱单一处理,说明共同胁迫对类囊体的堆叠造成的损伤可能较大,并且伤害程度大于单一处理。而单一处理与对照间没有显著差异。
植物叶片中类胡萝卜素可以吸收除长波和短波以外的其他剩余波长的光,耗散叶绿素吸收的过多光能,也是内源抗氧化剂。它除在光合作用中具有一定功能外,在细胞内还可以吸收剩余能,淬灭活性氧,从而防止膜质过氧化(Maslenkora et al., 1993)。图 4显示,与叶绿素a含量相似,处理前9天各处理间类胡萝卜素变化平缓,含量略高于或等于对照,说明轻度的胁迫能刺激类胡萝卜素的合成;第9~15天各处理急剧升高,盐处理升高的幅度最大,其次是干旱处理,协同处理升高幅度最小;之后盐处理呈较明显的下降趋势,其余处理变化平缓,且均低于对照,协同处理处于最低水平;复水后干旱处理的类胡萝卜素上升,而其他处理保持下降或平稳趋势,说明渗透胁迫对类胡萝卜素造成的伤害容易修复,而离子伤害的修复需要更长时间。

|
图 4 逆境胁迫对类胡萝卜素、类胡萝卜素含量与叶绿素a含量比率的影响 Figure 4 Effect of the stress on carotenoid content and ratio between carotenoid and chlorophyll a content |
类胡萝卜素含量与叶绿素含量比率的变化既与植物忍受逆境的能力有关,也能反映叶片光合活性的强弱(Willekens et al., 1994)。图 4还显示,在整个处理期间,盐和干旱单一处理呈现先降低后升高再降低的趋势,与对照的变化趋势相同,没有明显差异;协同处理在处理前12天与对照变化相反,以后呈降低的趋势,并且明显低于其他处理,这也间接说明协同处理对类胡萝卜素和叶绿素含量影响最大。
2.4 脯氨酸含量图 5显示,与对照相比,各处理的脯氨酸含量呈先升高后降低的趋势,只是上升与降低的时间不同,但均高于对照。前期各处理脯氨酸变化比较平缓,各处理之间没有明显差异;中期各处理间差异较大,盐与干旱协同处理的变化幅度最大,其次是干旱处理;复水后各处理均呈上升趋势,协同胁迫上升幅度最大。处理第6天,盐处理、干旱处理和盐与干旱协同处理的脯氨酸含量分别是对照的1.06,0.98和1.59倍,处理第15天分别是对照的1.62,5.76和9.4倍,处理第25天分别是对照的1.58,2.99和2.79倍,说明作为一种渗透调节物质,脯氨酸在处理的中、后期对维持细胞膜的稳定性、防护质膜透性方面发挥了重要作用,特别是在处理中期。

|
图 5 逆境胁迫对脯氨酸含量的影响 Figure 5 Effect of the stress on proline content |
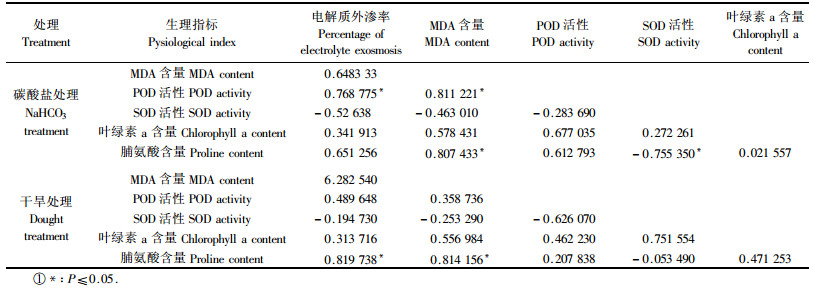
表 1显示了不同逆境胁迫下欧李几个生理指标之间的相关性。在碱盐胁迫下,欧李的电解质外渗率、MDA含量与POD活性间存在正相关关系,相关系数大多在0.6~0.8之间,以上3个指标与SOD活性之间存在一定程度的负相关;脯氨酸含量与电解质外渗率、MDA含量和POD活性间也存在正相关关系,而与SOD活性之间存在显著的负相关。干旱胁迫下,电解质外渗率、MDA含量与POD活性间存在着一定程度的正相关,但都没有达到显著水平;与碱盐胁迫相同,以上3个指标与SOD活性之间存在一定程度的负相关,图 2也显示胁迫处理前期POD活性增强,而处理后期SOD活性增强,说明POD活性与SOD活性具有一定的协同作用,但是他们起作用的时间存在差异;脯氨酸含量与电解质外渗率和MDA含量间存在显著的正相关,而与SOD活性之间存在微弱的负相关;说明随着胁迫程度的不断加深,脯氨酸作为一种渗透调节物质在维护细胞渗透势方面发挥了越来越强的作用。由此可见,欧李在碱盐处理和干旱处理条件下具有相似的生理反应,这也从侧面解释出碱盐胁迫会不同程度地造成植物的生理干旱,因此它们可能具有相似的生理机制。
|
|
在干旱与盐协同处理下,欧李通过以下途径来抵抗逆境:1)增强保护酶活性碳酸盐、干旱处理3 d后,各处理的POD活性明显高于对照;2)加快类胡萝卜素合成胁迫前期各处理的类胡萝卜素含量高于对照;3)增加渗透调节物质脯氨酸含量盐处理、干旱处理和盐与干旱共同处理的第15天脯氨酸含量分别是对照的1.62,5.76和9.4倍。
研究表明单一的碱盐处理不能对欧李的膜系统、酶系统和叶绿体造成伤害,但是单一的干旱处理能对其造成致命的伤害,可见与耐旱能力相比欧李具有较强的耐盐能力。但欧李在25 d的300 mmol·L-1 NaHCO3处理下其叶绿素a和类胡萝卜素含量呈明显下降趋势,这说明欧李还是受到了一定程度的伤害。有研究表明叶绿素是类囊体膜上色素蛋白复合体的重要组成,在盐胁迫下,植物叶片细胞中叶绿素与叶绿体蛋白间结合变得松弛,使更多的叶绿素遭到破坏,叶绿素含量下降(夏阳等,2005)。尽管本研究表明欧李可能具有较强的耐盐碱性,但由于处理时间有限,它是否能够耐受长期的高盐碱胁迫还有待进一步研究。
奥小平. 2006. 欧李的生态特性与栽培技术. 山西林业科技, 2006(1): 7-9. DOI:10.3969/j.issn.1007-726X.2006.01.003 |
楮建平, 孟平. 2008. 土壤水分胁迫对欧李幼苗光合及叶绿素荧光特性的影响. 林业科学研究, 21(3): 295-300. DOI:10.3321/j.issn:1001-1498.2008.03.003 |
郝再彬, 苍晶, 徐伸, 等. 2004. 植物生理实验. 2版. 哈尔滨: 哈尔滨工业大学出版社.
|
李志国, 王有信, 王玉峰, 等. 2005. 欧李加工品种与贮藏加工利用. 农产品加工, 2005(3): 21-24. DOI:10.3969/j.issn.1671-9646-B.2005.03.006 |
马建军, 张立彬, 刘玉艳, 等. 2008. 野生欧李生长期组织器官中不同形态钙含量的变化及其相关性. 园艺学报, 35(5): 631-636. DOI:10.3321/j.issn:0513-353X.2008.05.002 |
王有信, 何为军, 贺润平. 2007. 欧李杂交育种研究进展与新品种选育概况. 山西果树, 2007(1): 40-41. DOI:10.3969/j.issn.1005-345X.2007.01.025 |
王有信, 何为军, 李向东, 等. 2005. 欧李种质资源分布及种群分类特性研究. 山西果树, 2005(1): 35-36. DOI:10.3969/j.issn.1005-345X.2005.01.021 |
夏阳, 孙明高, 李国雷, 等. 2005. 盐胁迫对四种园林绿化树种叶片中叶绿素含量动态变化的影响. 山东农业大学学报, 36(1): 30-34. DOI:10.3969/j.issn.1000-2324.2005.01.006 |
姚岘武, 兰颜平, 周家华, 等. 2007. 欧李嫩枝扦插育苗及栽培技术. 林业实用技术, 2007(12): 27-28. |
张毅, 史传铎. 2003. 欧李的生物学特性与抗旱机理. 北京农业, 2003(9): 10-11. DOI:10.3969/j.issn.1000-6966.2003.09.017 |
张志良, 瞿伟菁. 2004. 植物生理学实验指导. 第3版. 北京: 高等教育出版社.
|
赵文恩, 韩雅珊, 乔旭光. 1999. 类胡萝卜素清除活性氧自由基的机理. 化学通报, (4): 1-4. |
庄丽娟, 苏福才. 2005. 欧李的组织培养与快速繁殖技术. 内蒙古农业大学学报, 26(1): 17-20. |
Maslenkova L T, Zanev Y, Popova L P. 1993. Adaptation to salinity as monitored by PSII oxygen evolving reactions in barley thylakoids. Plant Physiology, 142: 629-634. DOI:10.1016/S0176-1617(11)80410-4 |
Willekens H, Vancamp W, van Montagu M, et al. 1994. Ozone, sulfur dioxide, and ozone ultraviolet-B have similar effect on m-RNA accumulation of antioxidant genes in Nicotiana plum baginifolia L. Plant Physiology, 106: 1007-1014. DOI:10.1104/pp.106.3.1007 |