 2009, Vol. 45
2009, Vol. 45文章信息
- 赵中华, 惠刚盈, 袁士云, 刘文桢, 王润喜.
- Zhao Zhonghua, Hui Gangying, Yuan Shiyun, Liu Wenzhen, Wang Runxi
- 小陇山锐齿栎天然林空间结构特征
- Spatial Structure Characteristic of Quercus aliena var. acuteserrata Natural Forest in Xiaolongshan
- 林业科学, 2009, 45(3): 1-6.
- Scientia Silvae Sinicae, 2009, 45(3): 1-6.
-
文章历史
- 收稿日期:2008-01-16
-
作者相关文章



2. 甘肃省小陇山林业实验局 天水 741020
2. Xiaolongshan Forest Experiment Bureau of Gansu Province Tianshui 741020
森林空间结构特征体现了树木在林地上的分布格局及其属性在空间上的排列方式,决定了树木之间的竞争势及其空间生态位,在很大程度上决定了林分的稳定性、发展方向和经营措施(惠刚盈等,2007),已经成为森林经营中的重要因素。涉及单木之间空间关系的林分空间结构及其空间关系的描述和解释已成为森林结构研究的焦点(Moeur,1993;汤孟平,2003)。
小陇山林区是我国西北地区重要的天然林区,在水源涵养、保持水土、维护地区生态平衡、提高环境质量、保护生物多样性以及林业生产等方面发挥着不可替代的作用。在以往对小陇山林区的研究中,对构成该地区的主要森林群落锐齿栎(Quercus aliena var. acuteserrata)群落的生物量、群落数量特征、群落结构、种间关系、生物多样性等方面的研究较多(索安宁等,2004;2005;巨天珍,1995),但小陇山林区锐齿栎天然林空间结构特征的研究很少。本研究分析小陇山林区不同干扰类型锐齿栎天然林的空间结构特征及其差异,从优化森林空间结构的角度,为经营、保护和恢复小陇山林区的天然林和次生林提供借鉴。
1 研究区及样地概况小陇山林区(104°22′—105°43′E, 33°30′—34°49N′)位于甘肃省东南部,地处秦岭西端,我国华中、华北、喜马拉雅、蒙新四大自然植被区系的交汇处,属暖温带向北亚热带过渡地带,兼有我国南北气候特点,大多数地域属暖温湿润-中温半湿润大陆性季风气候。特殊的地理位置与环境条件加上生物的地理成分与区系成分复杂多样,使小陇山林区成为甘肃省生物种质资源最丰富的地区之一。小陇山林区海拔2 200 m以下主要分布有以锐齿栎和辽东栎(Quercus liaotungensis)为主的天然林,在栎林带内分布有华山松(Pinus armandi)、油松(Pinus tabulaeformis)、山杨(Populus davidiana)、漆树(Toxicodendron verniciflum)、冬瓜杨(Populus purdomii)、千金榆(Carpinus cordata)、甘肃山楂(Crataegus kansuensis)和刺楸(Kalopanax septemlobus)等乔木树种,灌木有美丽胡枝子(Lespedeza thunbergii)、中华绣线菊(Spiraea chinensis)、胡颓子(Elaeagnus pungens)、华北绣线菊(Spiraea fritschiana)、连翘(Forsytia suspense)、卫矛(Euonymus alatus)和山豆花(Lespedeza tomentosa)等。
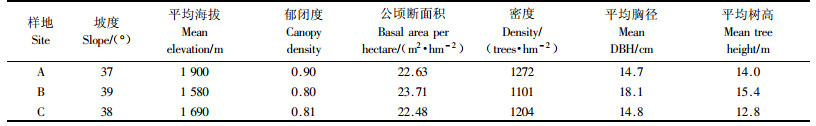
在小陇山林区百花林场王安沟营林区91林班14号小班(小班面积17 hm2,海拔1 900 m,坡向东北,平均坡度37°)选择100多年未经人为干扰的60 m×60 m的锐齿栎天然林为样地A,样地内的主要树种为锐齿栎,株数比例为61.5%,其他树种有膀胱果(Staphylea holocarpa)、青榨槭(Acer davidii)、漆树、泡花树(Craibiodendron stellatum)、三桠乌药(Lindera obtusiloba)等37个树种。样地设在山地深处,地理位置较偏僻,周围2 km范围内没有取材道,没有发现人为采伐痕迹。样地内有枯立木35株,胸径20 cm以上的有11株,20 cm以下的有24株;枯倒木162株,胸径20 cm以上的有93株,20 cm以下的有69株。树木最大年龄在110年以上。
将小陇山林业实验局李子园林场李子营林区的76林班18小班(小地名为响潭沟)和白营林区的159林班5小班(小地名为白营西沟)分别设为样地B和C:响潭沟为1969年皆伐利用锐齿栎天然林后天然更新形成的次生林,林内主要树种为锐齿栎,株数比例为63%,其他树种有山榆(Ulmus glabra)、网脉椴(Tilia paucicostata var.dictyoneura)、水榆花楸(Sorbus alnifolia)、油松等29个树种;白营西沟为1980和1995年对锐齿栎天然林进行2次大强度(30%)择伐利用后形成的林分,林内主要树种为锐齿栎,株数比例为79%,其他树种包括漆树、华山松、稠李(Padus racemosa)、油松等20个树种。小面积皆伐和大强度择伐是小陇山锐齿栎林的常见利用方式。A, B与C样地的概况见表 1。
|
|
对多年未经人为干扰的固定样地A运用TOPCON全站仪对胸径75 cm的植株进行每木定位,并做全面调查;对人为干扰样地B与C进行抽样调查。3块样的调查内容包括郁闭度、断面积、坡度、林分平均高、树种、胸径及其结构参数。树种、胸径及空间结构参数调查采用点抽样的方法,即从一个随机点开始,每隔一定距离(以调查的参照树的最近4株相邻木不重复为原则)设立一个抽样点,以激光判角器作为辅助设备,调查距抽样点最近的4株胸径大于5 cm树的树种、胸径级、角尺度、大小比数和混交度等空间结构参数,同时调查参照树与相邻树构成的结构单元的树种数,每个样地抽样点为49个,每个抽样点涉及4株参照树,每个参照树涉及4株相邻木。计算机模拟验证、东北红松(Pinus koraiensis)阔叶林样地验证以及厄瓜多尔天然林样地验证结果均表明,49个点作为天然林林木空间分布格局调查的最小样本量是合理的(徐海等,2007;惠刚盈等,2007)。
2.2 内业分析直径分布是林分内各种大小直径林木的分配状态,将直接影响到树木的树高、干形、材积、材种、树冠等因子(孟宪宇,1996)。本研究起测径为5 cm,径阶宽度为4 cm。
角尺度通过参照树与其最近4株相邻木构成的夹角是否大于标准角来描述相邻木围绕参照树的均匀性,不需要精密测距就可获得林木的水平分布格局,角尺度的取值有5种可能:0.00,0.25,0.50,0.75和1.00,林分平均角尺度落在[0.475,0.517]范围内,说明林木整体分布格局属随机分布,林分平均角尺度>0.517时林木为团状分布,林分平均角尺度<0.475时为均匀分布(Hui et al., 2002);
混交度用来说明混交林中树种的空间隔离程度(von Gadow, 1993;惠刚盈等,2001),混交度也有5种取值:0.00,0.25,0.50,0.75和1.00,分别描述为零度、弱度、中度、强度和极强度混交。修正林分混交度均值的M′计算公式为(惠刚盈等, 2007):
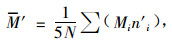
|
(1) |
式中:N为调查的林木株数;Mi为第i株树的混交度;n′i为第i株树所处的结构单元中的树种个数。
大小比数用来描述林木的大小分化程度(惠刚盈等,1999), 它量化了参照树与其相邻木的大小(胸径、树高或树冠等)相对关系。大小比数的5种取值为0.00,0.25,0.50,0.75和1.00,分别描述为优势、亚优势、中庸、劣态和绝对劣态,它明确定义了被分析的参照树在该结构块中所处的生态位,且其生态位的高低以中庸级为岭脊,生物意义十分明显。选取了3个林分中共有的树种:锐齿栎、华山松、色木槭(Acer mono)、漆树、白桦(Betula platyphylla)、水榆花楸、青榨槭、三桠乌药和樱桃(Cerasus polytricha),以胸径为指标对林分内林木大小的差异程度进行分析比较。
运用Winkelmass空间结构分析软件计算固定样地的结构参数,为避免边缘效应对林分结构的影响,Winkelmass在计算三个空间结构参数时设置了5 m缓冲区。
运用经典的K-S检验方法(Kolmogoroff,1933;Smirnoff,1939)进行不同林分类型的结构参数差异显著性检验。其原理是计算2个待检验抽样的相对累计曲线以及2者之间的最大绝对差异,以该最大绝对差异为检验变量与临界值进行比较,最大绝对差异计算公式为:
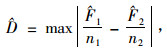
|
(2) |
式中: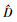
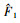
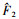
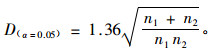
|
(3) |
如果 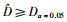
图 1表明,样地A百年未经干扰的锐齿栎天然林中小径阶的林木占多数,随着直径增大,林木株数减少;样地B近40年封育后自然恢复的天然林小径阶林木占多数,随着直径增大,林木株数急剧减少;样地C择伐林中虽然小径木占的比重也比较大,但各径阶间的减小幅度较为平缓,在林分中没有胸径为36~44 cm的林木,而有个别大于46 cm的“霸王树”。对3个林分的直径分布进行K-S差异显著性检验,结果表明:样地A与样地B直径分布差异不显著 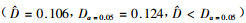
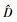

|
图 1 林分直径分布 Figure 1 Diameter distribution of stands |
对3个林分的角尺度(图 2)进行K-S差异显著性检验表明,3个林分的角尺度分布的差异不显著。样地C和A的角尺度均值分别为0.487和0.489,落在了[0.475,0.517]之间,属于随机分布的范畴;样地B锐齿栎次生林的平均角尺度为0.520,大于0.517,表现为轻微的团状分布;样地C林分原本为天然林,虽经历2次择伐但对其整体水平结构的改变不大,这是由于择伐本身伐除的是处于均匀或随机分布的大径木,所以其林分分布格局仍为随机分布,可以认为,择伐减少了聚集性,而增加了均匀性;王安沟锐齿栎天然林中林木个体总体上处于随机分布的状态,处于很均匀和团状分布这2种极端状况的情况较少,可以认为是群落长期自然演替的结果;响潭沟林分为天然林皆伐后天然更新的林分,至今尚未进行抚育,人为干扰较少,林分的形成可认为是自然演替的结果,林木有轻度聚集分布的现象,这也符合天然林演替过程中,林木分布的空间格局从聚集到随机的演替规律(孙冰等,1994;张家城等, 1999)。

|
图 2 林分角尺度及其频率分布 Figure 2 Uniform angle index and it's frequency distribution of stands |
图 3表明,样地A与B的林木混交度从0到1的相对频率逐渐增大,林分处于强度混交和极强度混交的比例分别达到73%和78.6%,2个林分中林木个体处于零度混交的比例分别为2.6%和0.6%,这说明林分中同种树聚集在一起的情况不多,大多数林木不与本种相伴;样地C林木总体上也是呈随混交度增大频率分布增大的趋势,但混交度从0.25到0.75的相对频率相差不大,而混交度为1的相对频率则达到38.8%,处于零度混交的相对频率也达到了9.6%。对3个林分的混交度进行K-S差异显著性检验,结果表明:样地A与B的混交度差异不显著(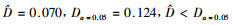


|
图 3 林分混交度及其频率分布 Figure 3 Mingling and it's frequency distribution of stands |
3个林分中共有的树种有9个,分析表明(图 4),在王安沟锐齿栎天然林林分中,白桦处于优势状态,水榆花楸、漆树、齿栎处和华山松处于亚优势地位并向中庸状态过渡,其他几个共有种则处于中庸或劣态;在响潭沟林分中,白桦、漆树、锐齿栎和青榨槭处于亚优势状态,色木槭、樱桃、三桠乌药和水榆花楸为中庸状态,而华山松处于劣态;白营西沟林分中,锐齿栎处于亚优势的状态,漆树则是亚优势向中庸状态过渡,白桦则是中庸状态,其他几个树种处于劣态或绝对劣态。3个林分共有种在林分中所处的地位有一定的差异,白桦在王安沟锐齿栎天然林、响潭沟林分和白营西沟林分的地位依次下降,即从处于优势地位到亚优势到中庸状态;华山松在王安沟林分中为亚优势向中庸状态过渡,而在另外2个林分中处于劣态;锐齿栎与漆树在3个林分中都处于亚优势向中庸过渡的状态;其他几个树种在3种林分中也表现出从王安沟锐齿栎天然林、响潭沟次生林、白营西沟次生林优势程度依次下降的趋势。这可能是由于白桦是群落演替的先锋种,在王安沟锐齿栎天然林群落中,经过长期的演替,白桦种群都以大径木的形式存在,而没有更新,即将退出群落,对另外2个林分而言,尚处于演替的早期,白桦还未占据优势地位;华山松随着演替进展,最终将作为顶极树种占据优势地位;锐齿栎和漆树等树种在林分中所处的地位与它们是构成当地森林群落的主要树种的事实是相符的,即在林分中处于各个径级的林木均有分布,更新良好,总体上表现为中庸的状态;此外,白营西沟林分中各树种所处的地位的重要原因可能是群落发育过程对人为干扰的响应,即抚育择伐改变了林分中各树种的地位。

|
图 4 3种林分共有树种平均大小比数 Figure 4 Mean value of neighborhood comparison of the mutual species in different stands |
森林生态系统,有其自身的发生和发展规律,森林经营必须遵循其规律(郑小贤,1999)。长期未经人为干扰的天然林群落能够按照自然规律向顶极群落演替,林分的直径结构特征呈负指数或负幂函数递减,林木的空间分布格局为随机分布,树种的隔离程度较高,且大小分化程度明显(Craig, 1980; Andrej, 2000;Linder et al., 1997;班勇等,1997;徐化成等, 1993; 1994;张家诚等,1999;臧润国等,2001),本研究中王安沟林分的结构具有明显的顶极群落特征。次生林可以理解为是原始森林生态系统的一种退化(Grau et al., 1997),次生林是我国森林资源的主体,是森林资源的重要基地。因此,次生林的经营是中国林业发展中最重要的主题之一(黄世能等,2000)。次生林经营就是人为地有目的地对次生林进行改造,从而使次生林的发展既符合自然演替规律,又满足人类的生态和经济效益。对次生林而言,如果任其自由发展,有可能顺向演替为原来的状态,这是植被自然趋同的结果,也是次生林经营的基础,但这一过程相当漫长(朱教君,2002),例如本研究中的响潭沟林分在皆伐迹地经过了长时间自然恢复, 虽然从林分的总体结构特征来说,是一个进展演替的过程,但林分林木的分布格局仍然呈轻微的团状分布,直径分布不尽合理,树种数较少,混交程度也不高。人为干扰活动对林分空间结构特征能产生重要的影响,例如白营西沟林分经历了2次大强度的择伐利用,林分的直径结构、树种隔离程度以及大小分化程度都发生了变化,与长期未经干扰的天然林及自然演替的次生林均存在显著差异。
小陇山林区是我国西北重要的天然林区,次生林为其主要的林分类型,合理地经营次生林是实现当地林业可持续发展的关键。因此,对小陇山次生林进行经营时,可根据生态学原理,从优化空间结构的角度出发,以长期未受干扰的天然林的结构特征为模板,调整林分的水平分布格局、树种组成和林内树种的竞争关系,使林分结构更接近于自然状态的林分结构特征,从而提升林分的整体功能。对响潭沟皆伐迹地恢复林分而言,通过抚育间伐,调整聚集分布的林木,减小相邻林木个体间的竞争,加速演替进程,使林木分布格局趋于随机。对白营西沟林分来说,可以通过补植、封育等措施来提高林分的混交度,减少人为活动的干扰,使林分结构尽快地恢复到合理状态。
班勇, 徐化成, 李湛东. 1997. 兴安落叶松老龄林落叶松林木死亡格局以及倒木对更新的影响. 应用生态学报, 8(5): 449-454. DOI:10.3321/j.issn:1001-9332.1997.05.001 |
黄世能, 王伯荪. 2000. 热带次生林群落动态研究:回顾与展望. 世界林业研究, 13(6): 7-13. DOI:10.3969/j.issn.1001-4241.2000.06.002 |
惠刚盈, 冯佳多克劳斯, 胡艳波, 等. 2007. 结构化森林经营. 北京: 中国林业出版社, 27-42.
|
惠刚盈, 胡艳波. 2001. 混交林树种空间隔离程度表达方式的研究. 林业科学研究, 14(1): 177-181. |
惠刚盈, GadowK, AlbertM. 1999. 一个新的林分空间结构参数——大小比数. 林业科学研究, 12(1): 1-6. DOI:10.3321/j.issn:1001-1498.1999.01.001 |
巨天珍. 1995. 天水小陇山锐齿栎群落种间关系定量分析. 西北植物学报, 15(3): 250-253. DOI:10.3321/j.issn:1000-4025.1995.03.015 |
孟宪宇. 1996. 测树学. 2版. 北京: 中国林业出版社, 66-80.
|
索安宁, 巨天珍, 张俊华, 等. 2004. 甘肃小陇山锐齿栎群落生物多样性特征分析. 西北植物学报, 24(10): 1877-1881. DOI:10.3321/j.issn:1000-4025.2004.10.019 |
索安宁, 巨天珍, 张俊华, 等. 2005. 甘肃小陇山锐齿栎群落生物量动态研究. 生态学杂志, 24(4): 377-381. DOI:10.3321/j.issn:1000-4890.2005.04.006 |
孙冰, 杨国亭, 迟福昌, 等. 1994. 白桦种群空间分布格局研究. 植物研究, 14(2): 201-207. |
汤孟平. 2003. 森林空间结构分析与优化经营模型的研究. 北京林业大学博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10022-2004021223.htm
|
徐海, 惠刚盈, 胡艳波, 等. 2007. 天然红松阔叶林林木分布格局研究的最小样本量. 林业科学研究, 20(2): 160-164. DOI:10.3321/j.issn:1001-1498.2007.02.002 |
徐化成, 范兆飞. 1993. 兴安落叶松原始林年龄结构动态的研究. 应用生态学报, 4(3): 229-233. DOI:10.3321/j.issn:1001-9332.1993.03.002 |
徐化成, 范兆飞, 王胜. 1994. 兴安落叶松原始林林木空间格局的研究. 生态学报, 14(2): 155-160. DOI:10.3321/j.issn:1000-0933.1994.02.009 |
于政中. 1993. 森林经理学. 北京: 中国林业出版社.
|
张家城, 陈力, 郭泉水, 等. 1999. 演替顶极阶段森林群落优势树种分布的变动趋势研究. 植物生态学报, 23(3): 256-268. DOI:10.3321/j.issn:1005-264X.1999.03.008 |
臧润国, 杨承彦, 蒋有绪. 2001. 海南岛霸王岭热带山地雨林群落结构及树种多样性特征的研究. 植物生态学报, 25(3): 270-275. DOI:10.3321/j.issn:1005-264X.2001.03.003 |
郑小贤. 1999. 森林资源经营管理. 北京: 中国林业出版社, 14: 89-92.
|
朱教君. 2002. 次生林经营基础研究进展. 应用生态学报, 13(12): 1689-1694. DOI:10.3321/j.issn:1001-9332.2002.12.040 |
Andrej B. 2000. Comparison of structure and biodiversity in the rajhenav virgin forest remnant and managed forest in the dinaric region of slovenia. Global Ecology and Biogeography, 9(3): 201-211. DOI:10.1046/j.1365-2699.2000.00155.x |
Craig G L. 1980. Age structure and disturbance history of a southern appalachian virgin forest. Ecology, 61(5): 1169-1184. DOI:10.2307/1936836 |
Grau H R, Arturi M F, Brown A D, et al. 1997. Floristic and structural patterns along a chronosequence of secondary forest succession in Argentinean subtropical montane forests. Forest and Ecology Management, 95: 161-171. DOI:10.1016/S0378-1127(97)00010-8 |
Hui G Y, von Gadow K. 2002. Das winkelmass-theoretische überlegungen zum optimalen standardwinkel. Allgemeine Forst und Jagdzeitung, 173(9): 173-177. |
Kolmogoroff A N. 1933. Sulla determinazione empirica di una legge di distribuzione. Giornale Instituto Italiano Attuari, 4: 441-452. |
Linder P, Elfving B, Zackrisson O. 1997. Stand structure and successional trends in virgin boreal forest reserves in Sweden. Forest Ecology and Management, 98(1): 17-33. DOI:10.1016/S0378-1127(97)00076-5 |
Moeur M. 1993. Characterizing spatial patterns of trees using stem mapped data. Forest Science, 39(4): 756-775. |
von Gadow K. 1993. Zur bestandesbeschreibung in der forsteinrichtung. Forest und Holz, 48(21): 602-606. |