 2009, Vol. 45
2009, Vol. 45文章信息
- Zhang Yanfeng, Xie Yingping, Xue Jiaoliang, Wang Xu
- 张艳峰, 谢映平, 薛皎亮, 王旭
- Attraction to the Ladybeetle by the Volatiles of Persimmon Trees Induced with Methyl Jasmonate and Japanese Wax Scale Attacking
- 茉莉酸甲酯和日本龟蜡蚧诱导柿树挥发物对红点唇瓢虫的吸引
- Scientia Silvae Sinicae, 2009, 45(1): 90-96.
- 林业科学, 2009, 45(1): 90-96.
-
文章历史
- 收稿日期:2008-09-08
-
作者相关文章



Persimmon (Diospyros kaki)is an important fruit in northern China where about 36 700 hm2 of persimmon orchards are planted. However, persimmon trees were often attacked by many scale insects, in which Japanese wax scale, Ceroplastes japonicus (Hemiptera: Coccoidea: Coccidae) is the serverest one. Because of infestation of the scale insects, 70% of fruit yields were reduced annually. Japanese wax scale secreted mass wax substances to form a thick wax layer covering on its body dorsal surface. The wax protected the scale insect from attacking by other factors including chemical pesticides. Therefore, it is very difficult to control the wax scale with pesticides (Xie, 1998). Applying natural enemy insects to control scale insects has been considered as a better strategy. In biological control, many researches were reported that when they were attacked by insect pests, plants could be induced to release some volatile chemicals that functions as a signal of prey to the pest's natural enemies (Turlings et al., 1991; 1995; 1998; Takeshi et al., 1997; Han et al., 2004). For example, Souissi et al. (1998; 1999) found that mealybug-infested plants were the major sources of volatiles that attracted female parasitoids to the microhabitat of its host. Xie et al. (2004) reported that the fresh twigs and leaves of bunge prickly-ash trees (Zanthoxylum bungeanus) damaged by a mealybug (Phenacoccus azaleae) showed an attractive effect to a ladybeetle (Harmonia axyridis), the predator of the mealybug.
Our previous study found, however, the persimmon tree damaged by Japanese wax scale did not show an equal attraction in different seasons to the predators, such as Chrysopa sinica (Neuroptera: Chrysopidae), Harmonia axyridis and Chilocorus kuwanae (Coleoptera: Coccinellidae). In July and early August, the attraction effect was less, but in September the attraction effect was strong (Yang et al., 2006). This resulted in only a few predators occurred in persimmon tree orchards and they could not effectively control the infestation of the scale insect during July and early August. In fact, in this period, persimmon fruit growth was at the rapid phase. Japanese wax scale nymphs were feeding on the leaves and young twigs. Their population often reached average 16 (max. 200) each leaf. So that fruit yields were significantly affected by the infestation of the wax scale. It is very important to increase the attractive capability of the persimmon trees to recruit more natural enemy insects to control the scale insects.
Recent years, methyl jasmonate (MeJA) and jasmonic acid (JA) were reported on inducing plant volatile to attract predators and parasitoids of pest insects (Rodriguez-Saona et al., 2001; Lou et al., 2006). Dicke (1999) found that Phytoseiulus persimilis, a carnivorous mite and predator of the two-spotted spider mite (Tetranychus urticae) a pest of lima bean plants was attracted by the volatiles from JA-treated lima bean plants. Degenhardt et al. (2006) examined the effect of herbivory and MeJA exposure on the release of volatile organic compounds (VOCs) in the marsh elder (Iva frutescens). They found that I. frutescens produced novel VOCs in response to feeding by the special leaf beetle (Paria aterrima) and MeJA treatment.
In this study, we investigated the potential capability of persimmon trees to produce attractive volatiles in response to exposure of MeJA in July. The ladybeetle (Chilocorus kuwanae), a primary predator of Japanese wax scale was chosen in this trail. Some questions that will be addressed are: 1) How fluctuated attractive ability of the persimmon trees damaged by Japanese wax scale nymphs around a whole day? 2) Could an exogenous MeJA application induce persimmon trees releasing more volatiles to attract the ladybeetles and what about the change fluctuation? 3) What are the chemical components worked as attractant in the volatiles?
1 Materials and methods 1.1 Experimental materials and setupThe study was conducted in two persimmon orchards located at Linyi County of Shanxi Province in northern China (35°40′N, 111°27′E, Elev. 700 m). Two-year-old persimmon trees about 1.5 m in height and 1.0 m in crown width were used in the study. A large population of Japanese wax scale was found in the damaged orchard. The average density was 16 (max. 200) on each leaf. The damaged orchard is about 1 km away from the natural undamaged one. The two orchards are on the same soil type and have the same climate.
Ladybeetles were collected from local persimmon tree orchards and their responses were tested to the volatiles from the persimmon trees induced by the wax scale infestation or by exogenous MeJA application with a series of experiments.
1) Experiment 1: We collected leaves from persimmon trees damaged by the wax scale nymphs and from healthy trees at 10:00, 15:00, 19:00, and 7:00 of the second day. Laboratory tests for ladybeetle response were conducted immediately following the collection.
2) Experiment 2: We applied 100 mL solution containing 200 μL of a 10:1(ethanol/MeJA) mixture to healthy trees. Leaves from treated and untreated trees were collected at 10:00, 15:00, 19:00, and 7:00 of the next day, which correspond to 3, 8, 12 and 24 h after MeJA was applied. Untreated trees in a separate site were sprayed with a solution without MeJA. Laboratory test for ladybeetle response was immediately following the collection.
3) Experiment 3: Based on the volatiles identified from trees that were treated with the MeJA, we selected two compounds, α-Pinene and Limonene, to test ladybeetle response in three consecutive days. We used 4 concentrations from 10-3 to 10-6 g·mL-1 for each compound (Han et al., 2004), 2 mL each time.
1.2 Test proceduresLadybeetle response to the persimmon volatiles was measured with an olfactometer, a Y-shaped glass tube consisted of the base and two arms. Each arm was connected to an odor source container, in which 10 pieces of intact leaf materials was used. An airstream was generated and led through a tube with active charcoal and a humidifier bottle. It was divided in two secondary airstream through the odor containers, then through the two arms of the Y-tube olfactometer at a rate of 200 mL·min-1. The Y-tube olfactometer was placed in plane with a 30 W artificial light source. During the experiments, the temperature was maintained at 25~28 ℃.
Persimmon leaves as the odor source were collected from the sample trees. 15 ladybeetles were introduced individually into the base tube of the olfactometer and given 10 mins to crawl towards the odor source. A positive tropism choice for an odor source was defined by a ladybeetle crawling 1/3 of the length of the Y arm and remaining there for at least 1 min. Ladybeetles that did not make a choice within 10 min were recorded as "no response." To eliminate the effects of asymmetrical bias, connections of the two arms of the olfactometer to the odor source containers were exchanged after testing of 15 ladybeetles. After 30 ladybeetles were tested, the olfactometer tube was washed and heated to dry and the odor source containers were exchanged. For each odor source group, the test was repeated 6 times per day for three consecutive days. In each replication, different female adult ladybeetles were used.
1.3 Volatile chemicals collection and analysisWe collected volatile chemicals on persimmon trees from plots treated with exogenous MeJA, and control. Volatiles were collected from all sample trees within 24 h after MeJA treatment to identify the variety of those compounds released and the relation with attraction to ladybeetles. According to previous study, control trees were also collected at 15:00 which possess the most volatile emission amounts. Then, we tested ladybeetle response to two compounds.
A headspace volatile trapping instrument was used to collect volatiles from persimmon trees. An air pump (model QC-1S) was used to pull and control airflow through the volatile-collection-bag (80 cm long, 45 cm in diameter). Air entered the bottom of the bag through an air-filtration-tube to keep the air clean and dry, passed through a volatile-absorption-tube filled with a sorbent, Tenax GR (60/80 mesh), and exited out the bag. In the volatile collection process, the airflow rate was controlled at 100 mL·min-1 through the volatile-collection-bag. The collection time for each volatile sample lasted for 1 hour and each treatment was repeated 5 times.
We analyzed chemical compositions of the volatile samples with a Thermal-Desorption Cold Trap-Gas Chromatograph/Mass Spectrometry (TCT-GC/MS). TCT (Chrompark) Model: CPG-4010PTI/TCT, system press: 20 kPa, temperature of the inlet and cold trap: 250 and -120℃. All samples were analyzed by GC/MS (TraceTM2000GC) equipped with a CP-Sil 8 Low Bleed/MS column (60 m × 0.32 m ID, 0.5 μm) with helium as a carrier gas. Following thermal desorption, column temperature was maintained at 40 ℃ for 3 min and increased at the rate 6 ℃·min-1 to 270 ℃ and maintained for 3 min, then increased to 280 ℃ for 5 min. The compounds of volatiles were identified by comparison of retention time and mass spectra to those contained in the standards database of NIST98 library (Scientific Instrument Services, Inc., USA).
1.4 Statistical analysesWe tested the differences in ladybeetle response to volatile sources between from the damaged or treated leaves and from healthy leaves across times in each experiment. We used Chi-square statistics to the test null hypothesis that ladybeetles respond to two volatile sources evenly. In addition, two-or three-way contingency tables with likelihood-ratio Chi-square (G2) were used to test the interactions between times when leaves were collected. The pair-wise comparisons among times were tested using the Bonferroni adjustment (P=0.05/numbers of pairs). Individual tests were performed for each of the volatiles collected from leaves. Significance level was set as α=0.05 for statistical analysis. Only probabilities that showed a significant difference were presented. The statistical analysis was performed either with PROC FREQ in SAS (V9.12, SAS Institute Inc.) or SPSS (V12.0, SPSS Inc.).
2 Results 2.1 Ladybeetle response to persimmon trees induced by wax scale infestation or MeJAThe results from the 4 time phases in each day across three testing dates in July showed that the attraction of persimmon trees infested by Japanese wax scale nymphs fluctuated a little (Fig. 1a). The strongest attractive effect was 56.76% which appeared at 15:00. In general, no significant difference was found in ladybeetle response between the two odor sources from nymph infested leaves and healthy leaves (χ2 < 1.83, df=3, P > 0.61). No interactions were found among odor sources, test dates, and time phases when leaves were collected (G2 < 3.84, df varies, P > 0.69). Based on 90 ladybeetles tested for each treatment, an average of 41 ladybeetles responded to the infested leaves and 35 responded to healthy leaves, respectively. In summary, Japanese wax scale nymphs feeding could not induce the persimmon trees to release enough volatile to attract ladybeetles, C. kuwanae.

|
Fig.1 Percentage of ladybeetle Chilocorus kuwanae response (+SD) to the odors released from healthy persimmon trees and (a) trees damaged by nymphs of Japanese wax scale in July and (b) trees treated with 20 μl Methyl jasmonate (MeJA) at 4 times during a day A total of 90 ladybeetles were tested at each time. The ladybeetles that did not respond were excluded from the analysis. The numbers embedded in the bars are the numbers of ladybeetles responded (**: P < 0.01) |
After persimmon trees were treated with MeJA, significantly more ladybeetles crawled toward the odor source from treated trees (Fig. 1b). The differences were significant for each of the three testing dates (χ2 > 8.52, df=3, P < 0.04). Three-way interaction was significant (G2=39.56, df=17, P > 0.01) and so was the interaction between odor source and time phases when leaves were collected (G2=34.46, df=3, P < 0.01). Both interactions were caused by leaf collection time phases across three testing dates. At the 15:00 collections, an average of 61 ladybeetles moved toward the MeJA treated leaves, but only 17 ladybeetles crawled toward the untreated leaves, so corresponding attractive effects were 78.21% and 21.79% (χ2 > 20.00, df=1, P < 0.01). A similar result was found in the 7:00 am next day collection and the attractive effects were 73.42% and 26.58% (χ2 > 11.31, df=1, P < 0.01). However, at the sampling time phases of 10:00 and 19:00 ladybeetle response was not significant difference between treated and untreated leaves.
2.2 Composition of the volatile emissionsIn July, when ladybeetles responded to the odors of MeJA-treatment persimmon trees, the volatiles were simultaneously collected from the treated trees and untreated trees. 17 chemical compounds were detected from the 2 classes of volatiles and they could be divided into 4 groups, including terpenoid (6), ester (2), hydrocarbon (8) and aldehyde (1) (Tab. 1). The first 2 groups were commonly considered as the most interest compounds in attraction to the natural enemy insects. The first group contained 6 terpenoids which only 4 were detected from the volatile of control trees, but other two, 2[10]-pinene and D-limonene as novel components were discovered in the volatiles of the MeJA-treated trees. Other 3 terpenoid components, e.g. 3-thujene, α-pinene, and 4[10]-thujene, became much more in quantity, especially, α-pinene reached 41.19% (P < 0.01). In contrast, Ocimene sharply decreased in relative amounts after being treated with MeJA. Meanwhile, the second group included two ester compounds. The relative quantity of 3-hexenyl-acetate became higher in the volatiles from the trees treated with MeJA than that form the control. But dibutyl phthalate was detected out from the volatiles of MeJA-treated trees only. Hydrocarbons, as the third group, accounted for a high ratio (68.87%) in the volatiles of control trees, whereas a low ratio (4.62%) in the volatiles of treated trees. The fourth group only one aldehyde compound, nonanal, appeared just in the volatiles of treated trees.
|
|

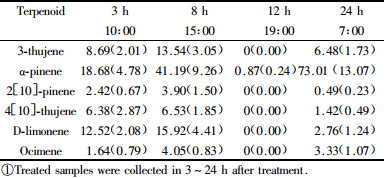
Based on terpenoid compounds fluctuation in relative quantity (Tab. 2), an interesting phenomenon was found. From 3 h to 8 h after MeJA treated the relative content of the 6 terpenoids all increased and the total quantity was 50.33% and 85.13% respectively, especially, 3-thujene and α-pinene increased obviously. However, up to 12 h that corresponding to 19:00, only α-pinene was detected in a very low content (0.87%). But to 7:00 in the following morning, the 6 terpenoids were detected again and the content of α-pinene even reached 73.01%.
|
|
Based on analysis of the chemical compositions of the volatiles from the trees treated with MeJA, two single chemical compounds were selected in trail for confirming their attractive effect to the ladybeetles, C. kuwanae (Fig. 2). We found that α-pinene and D-limonene displayed a significant attraction to the ladybeetles (χ2 >8.48, df=3, P < 0.04). The attraction effect of α-pinene appeared in the concentration range from 1×10-4 to 1×10-6 g·mL-1 and the strongest attraction was 75.61% at 1×10-5 g·mL-1. D-limonene at the concentrations of 1×10-4 and 1×10-5 g·mL-1 showed obvious attraction to ladybeetles, while the stronger attraction was 78.95% at 1×10-4 g·mL-1.

|
Fig.2 Percentage of ladybeetle Chilocorus kuwanae response to the odors of (a) α-pinene and (b) D-limonene at 4 concentrations A total of 90 ladybeetles were tested at each concentration. The ladybeetles that did not respond were excluded from the analysis. The numbers embedded in the bars are the numbers of ladybeetles responded (**:P < 0.01 and *:P < 0.05) |
It has been found that herbivory induce host plants to release volatile chemicals, which subsequently attract natural enemies to control pests (Turling et al., 1990, Dick et al., 1993; Kessler et al., 2001). However, our study found that persimmon trees under attacking by Japanese wax scale did not always display equally attractive effect to predators, Chrysopa sinica, Harmonia axyridis and Chilocorus kuwanae. In July, Japanese wax scale nymphs fed on host leaves and twigs and the damaged trees demonstrated invisible attractive effect to the natural enemies. Maybe the population of the wax scale does not enough to turn on the switch of persimmon trees on defence. But in September, the wax scale adult fed heavily with the average density was 32 (max. 240) per 10 cm on twigs. And the damaged trees produced a strong attractiveness (Yang et al., 2006). This pattern suggests that host plant releasing attractive volatiles to natural enemies correlates not only with the herbivorous attack but also with herbivorous developmental stages, population density or probably host plant physiology and phenology.
In present study, we found the attraction of persimmon trees infested by Japanese wax scale nymphs fluctuated in different time phases around a whole day (Fig. 1a). Although the strongest attractive effect appeared at 15:00, no significant difference was found in ladybeetle response between the two odor sources from nymph infested leaves and healthy leaves. MeJA application in July made much more ladybeetles crawled toward the odor source from treated trees (Fig. 1b). It means applying MeJA in July could induce persimmon trees to change their volatile emission to attract more ladybeetles. Thaler (1999) reported inducing tomato plants with jasmonic acid increased parasitism of caterpillar pests in an agricultural field twofold. Rodriguez-Saona et al. (2001) reported exogenous Methyl jasmonate could induce cotton plants to produce similar volatiles with those produced from plants attacked by beet armyworm, Spodoptera exigua. Comparatively, attractiveness fluctuation of volatiles of persimmon trees during a day was noted rarely in the past.
The chemical composition analysis of the volatiles revealed the reason that the predators prefer to aggregate to the odor sources of the persimmon trees treated with MeJA. In the volatiles of MeJA treated trees, 6 novel components appeared including 2 terpenoids, 2[10]-pinene and D-limonene. Simultaneously 4 compounds increased their relative contents including 3 terpenoids, especially α-pinene (Tab. 1). Furthermore comparing with the ladybeetle response, more than 85% terpenoids were the most primary components in the attractive volatiles of persimmon trees. In which α-pinene, 3-thujene and limonene were the dominant ones. It is similar with others reports that terpenoids were the main attractants for natural enemies (Dicke et al., 1999). However, in 4 time phases 19:00 showed a significantly different with other phases, only α-pinene was detected in a very low content (0.87%). Probably, it is affected by light because many of volatile compounds (especially terpenoids) are strongly regulated by light, which may be a major factor influencing emissions and subsequent attraction of natural enemies during this sampling period. Since the terpenoids of volatiles played an important role in attracting natural enemies, we used two terpenoids which are available in the market, α-pinene and limonene, for the test. The results suggest that those two compounds were highly attractive to the predator, C. kuwanae, at the specific concentrations.
In conclusion, Methyl jasmonate is a signal transducer to effectively induce plants to change volatiles emission in component and quantity. Therefore, it is interest in biocontrol by applying MeJA in orchards to induce persimmon trees to emit attractive volatiles to recruit ladybeetles in July, the stage of Japanese wax scale nymphs infestation.
Acknowledgements
We thank Drs. Zhang Jianwei and William Oliver (USDA Forest Service, Pacific Southwest Research Station, 3644 Avtech Parkway, Redding, California 96002) who not only edited our English for this manuscript, but also provided many thoughtful comments. The help in statistical analysis from Dr. Zhang is greatly appreciated.
Degenhardt D C, Lincoln D E. 2006. Volatile emissions from an odorous plant in response to herbivory and methyl jasmonate exposure. J Chem Ecol, 32: 725-743. DOI:10.1007/s10886-006-9030-2 |
Dick M, Baarlen P V, Wessels R, et al. 1993. Herbivory induces systemic production of plant volatiles that attract predators of the herbivore: Extraction of endogenous elicitor. J Chem Ecol, 19: 581-599. DOI:10.1007/BF00994327 |
Dicke M, Gols R, Ludeking D, et al. 1999. Jasmonic acid and herbivory differentially induces carnivore-attracting plant volatiles in lima bean plants. J Chem Ecol, 25: 1907-1922. DOI:10.1023/A:1020942102181 |
Dicke M.1999. Evolution of induced indirect defense of plants//Tollrian R, Harvell C D. The Ecology and Evolution of Inducible Defenses. Princeton University Press, Princeton, New Jersey, 483-520.
|
Han Baoyu (韩宝瑜), Chen Zongmao (陈宗懋).2000. Behavior response of four Leis axyridis varieties to volatiles from tea and Toxoptera aurantii. Chin J Appl Ecol (应用生态学报), 11: 413-416. http://en.cnki.com.cn/Article_en/CJFDTOTAL-YYSB200003020.htm
|
Han Baoyu (韩宝瑜), Chen Zongmao (陈宗懋).2004. Attraction effect of main volatile components from tea shoots and flowers on Sphaerophoria menthastri (Diptera: Syrphidae) and Chrysopa septempunctata (Neuroptera: Chrysopidae). Chin J Appl Ecol (应用生态学报), 15(4): 623-626. http://en.cnki.com.cn/Article_en/CJFDTOTAL-YYSB200404016.htm
|
Hare J D, Walling L L. 2006. Constitutive and Jasmonate-inducible traits of Datura wrightii. J Chem Ecol, 32: 29-47. DOI:10.1007/s10886-006-9349-8 |
Kessler A, Baldwin I T. 2001. Defensive function of herbivory-induced plant volatile emissions in nature. Science, 291: 2141-2214. DOI:10.1126/science.291.5511.2141 |
Lou Yonggen, Hua Xiaoyan, Turlings T C J, et al. 2006. Differences in induced volatile emission among rice varieties result in differential attraction and parasitism of Nilaparvata lugens eggs by parasitoid Anagrus nilaparvatae in the field. J Chem Ecol, 32: 2375-2387. DOI:10.1007/s10886-006-9151-7 |
Rodriguez-Saona C, Crafts-Brandner S J, Pare R W, et al. 2001. Exogenous methyl jasmonate induces volatile emissions in cotton plants. J Chem Ecol, 27: 679-695. DOI:10.1023/A:1010393700918 |
Souissi R, Nenon J P, Le R B. 1998. Olfactory responses of parasitoid Apoanagyrus lopezi to odor of plants, mealybugs, and plant-mealybug complexes. J Chem Ecol, 24: 37-38. DOI:10.1023/A:1022332711331 |
Souissi R, Le R B. 1999. Behavioral responses of the endoparasitoid Apoanagytus lopezi to odours of the host and host's cassava plants. Ent Exp Appl, 90: 215-220. DOI:10.1046/j.1570-7458.1999.00441.x |
Takeshi S, Takabayashi J, Ashihara W, et al. 1997. Response of predatory insect Scolothips takahashii toward herbivore-induced plant volatiles under laboratory and field condition. J Chem Ecol, 23: 2033-2048. DOI:10.1023/B:JOEC.0000006487.49221.df |
Thaler J S. 1999. Jasmonate-inducible plant defences cause increased parasitism of herbivores. Nature, 399: 686-688. DOI:10.1038/21420 |
Turlings T C J, Benrey B. 1998. Effect of plant metabolites on the behavior and development of parastic wasps. EcoScience, 5: 321-333. DOI:10.1080/11956860.1998.11682472 |
Turlings T C J, Tumlinson J H, Heath R R, et al. 1991. Isolation and identification of allelochemicals that attract the larval parasitoid, Cotesia marginiventris (Cresson), to the microhabitat of one of its hosts. J Chem Ecol, 17: 2235-2251. DOI:10.1007/BF00988004 |
Turlings T C J, Tumlinson J H, Lewis W J. 1990. Exploitation of herbivore-induced plant odors by host seeking parasitic wasps. Science, 250: 1251-1253. DOI:10.1126/science.250.4985.1251 |
Turlings T C J, Loughrin J H, Mccall P J, et al. 1995. How caterpillar-damaged plants protect themselves by attracting parasitic wasps. Proc Natl Acad Sci USA, 92: 4169-4174. DOI:10.1073/pnas.92.10.4169 |
Turlings T C J, Lengwiler U, Bernasconi M L, et al. 1998. Timing of induced volatile emissions in maize seedlings. Planta, 207: 146-152. DOI:10.1007/s004250050466 |
Xie Yingping (谢映平).1998. The Scale Insects of the Forest and Fruit Trees in Shanxi of China. Beijing: China Forestry Publishing House, 22-38.
|
Xie Yingping (谢映平), Xue Jiaoliang (薛皎亮), Tang Xiaoyan (唐晓燕), et al. 2004. The bunge prickly-ash tree damaged by a mealybug, Phenacoccus azaleae attracting the ladybug, Harmonia axyridis. Scientia Silvae Sinicae(林业科学), 40(5): 116-122. http://www.linyekexue.net/CN/abstract/abstract4059.shtml
|
Yang, Xingen (杨新根), Xie Yingping (谢映平), Xue Jiaoliang (薛皎亮), et al. 2006. Change in volatiles of Diospyros kaki damaged by Ceroplastes japonicus Green and their attraction to Chilocorus kuwanae Silvestri. Chin J Appl Environ Biol (应用与环境生物学报), 12: 215-219.
|