 2009, Vol. 45
2009, Vol. 45文章信息
- 张蕊, 周志春, 金国庆, 骆文坚.
- Zhang Rui, Zhou Zhichun, Jin Guoqing, Luo Wenjian
- 南方红豆杉种源遗传多样性和遗传分化
- Genetic Diversity and Genetic Differentiation of Taxus wallichiana var. mairei Provenances
- 林业科学, 2009, 45(1): 50-56.
- Scientia Silvae Sinicae, 2009, 45(1): 50-56.
-
文章历史
- 收稿日期:2008-01-30
-
作者相关文章



2. 浙江省林业种苗管理总站 杭州 310020
2. Forestry Seed Administration of Zhejiang Province Hangzhou 310020
红豆杉属(Taxus)植物是一类古老的植物类群,全世界有11种,广泛分布于欧洲、北美洲及东亚等北半球地区,我国有3种2变种,即东北红豆杉(T. cuspidata)、喜马拉雅密叶红豆杉(T. funana)、喜马拉雅红豆杉(T. wallichiana)和2变种红豆杉(T. wallichiana var. chinensis)、南方红豆杉(T. wallichiana var. mairei)(Fu et al., 1999)。红豆杉属植物集药用、材用和观赏等于一体,开发利用价值极高,尤其自20世纪70年代从短叶红豆杉(T. brevifolia)树皮中提取紫杉醇(taxol)以来,该属植物倍受人们重视,但这给红豆杉属野生资源带来了毁灭性的破坏,使其处于濒危状态。为保护红豆杉属自然资源免遭绝灭,许多国家已明令禁止采伐,红豆杉属植物及其制品的国际贸易受到严格控制。为实现红豆杉属植物的有效保护和利用,各国皆加强了遗传多样性等研究,以制定科学的遗传保育策略。如Lewandowski等(1995)利用18个同工酶位点研究发现波兰地区欧洲红豆杉(T. baccata)种群具有较高的遗传多样性,可提供有较高利用和保护价值的优良种质。Senneville等(2001)利用等位酶研究表明,加拿大魁北克地区6个加拿大红豆杉(T. canadensis)天然种群的遗传多样性很低,种群遗传分化很高,认为是由于种子传播方式使其迁徙扩张受到限制之故。Shah等(2007)利用RAPD分子标记研究发现巴基斯坦地区密叶红豆杉(T. fauna)的遗传多样性很低,并因其生境严重的片断化及缺乏有效的基因流而导致种群间产生巨大的遗传分化。我国学者Li等(2006)利用RAPD分子标记研究证实了东北红豆杉一个变种(T. cuspidata var. nana)存在很高的遗传多样性。
南方红豆杉是中国红豆杉属植物中分布最为广泛的一个种,自然分布于亚热带各省区。南方红豆杉材质优异,是高档珍贵用材,其紫杉醇含量虽稍低于喜马拉雅红豆杉等,但因早期速生、生物收获量大、适宜短周期作业而具有巨大的药用开发利用价值。通过多年的研究,我国已突破了南方红豆杉短周期药用林培育的技术关键,实现了栽植2~3年就能收获利用的目标。通过10省区27个产地的苗期地理种源试验,揭示了南方红豆杉苗木生长的种源变异规律(焦月玲等, 2007)。南方红豆杉属国家一级保护濒危树种,但有关致濒机制和遗传多样性等相关研究较少。周其兴等(1998)利用等位酶技术研究了广西、湖南和贵州3个自然种群及同属其他植物的遗传多样性,发现我国红豆杉属植物的遗传多样性水平较高,国产的3种1变种的遗传一致度较高。张宏意等(2003)基于RAPD分子标记对广东、湖南和江西较小分布区12个自然种群的遗传多样性进行研究,结果发现地域相距较远种群间的遗传分化较大,粤北种群的遗传多样性较低。本文在南方红豆杉全分布区地理种源试验的基础上(焦月玲等, 2007),选择10个省区15个代表性种源,利用ISSR分子标记研究南方红豆杉整个分布区种源的遗传多样性及地理变化、种源遗传分化等,从而为科学制定南方红豆杉遗传保育策略提供理论依据。
1 材料与方法 1.1 植物样品采集试验叶样取自浙江省淳安县富溪林场圃地的南方红豆杉种源试验2年生留床苗。取样种源来自10个省区的15个代表性种源,包括安徽黄山,浙江龙泉,福建武夷山、明溪、武平,江西庐山、井冈山,湖南桑植、绥宁,湖北恩施,贵州黎平、都匀,四川峨眉,广西三江,云南石屏。南方红豆杉种源试验的采种林分皆为当地起源的天然林,除福建武平和武夷山等较小种群外,要求在采种天然林分中选择20株以上优良母树采种,母树间距50 m以上,每株母树等量采种、混合处理后作为该种源的种子,每个种源提供种子2.5 kg。参试种源基本情况和苗期遗传测定等详见焦月玲等(2007)一文。2007年7月中旬,从上述15个种源中每种源随机选取20株单株,分别采集其顶端新发枝条上的新鲜嫩叶,将其放入装有生物冰袋的保温箱中带回实验室,置于-20 ℃冰箱中保存。

|
图 1 15个南方红豆杉试验种源的分布示意图 Figure 1 Distribution for 15 tested provenances of T. wallichiana var. mairei 1.安徽黄山Huangshan, Anhui; 2.浙江龙泉Longquan, Zhejiang; 3.福建武夷山Wuyishan, Fujian; 4.福建明溪Mingxi, Fujian; 5.福建武平Wuping, Fujian; 6.江西庐山Lushan, Jiangxi; 7.江西井冈山Jinggangshan, Jiangxi; 8.湖南桑植Sangzhi, Hunan; 9.湖南绥宁Suining, Hunan; 10.湖北恩施Enshi, Hubei; 11.贵州黎平Liping, Guizhou; 12.贵州都匀Duyun, Guizhou;13.四川峨眉Emei, Sichuan; 14.广西三江Sanjiang, Guangxi; 15.云南石屏Shiping, Yunnan |
每株取0.5 g新鲜嫩叶,采用改良CTAB法提取基因组DNA(Doyle et al., 1987),提取的DNA溶于1×TE缓冲液中,用1.0%的琼脂糖凝胶电泳检测其质量,并在紫外分光光度计(日本岛津UV-2401PC)下检测其浓度,最后稀释至40 ng·μL-1,放入-20 ℃冰箱储存备用。
1.3 PCR扩增PCR扩增反应在PTC-100TM基因扩增仪上进行。所用引物为加拿大哥伦比亚大学UBC公司公布的第9套ISSR引物,从50个ISSR引物中筛选出扩增条带清晰、反应稳定的8个引物对所有供试植株的DNA样品进行扩增。PCR扩增体系20 μL,包括:Taq酶1.00 U、dNTP 0.20 mmol·L-1、MgCl2 1.0 mmol·L-1、引物0.3 μmol·L-1、基因组DNA 80 ng。PCR反应程序为:94 ℃ 4 min;94 ℃ 45 s,44~58 ℃ 45 s(引物退火温度随引物的碱基含量变化略作调整,见表 1),72 ℃ 1.5 min,35个循环;72 ℃延伸5 min,16 ℃下保存。PCR产物在1.5%的琼脂糖凝胶电泳分离,对照标准分子量DGL 2000 DNA Marker(北京鼎国生物技术有限责任公司),再经Gel Red荧光染料染色,最后利用复日FR-200紫外与可见光分析成像系统拍照记录结果。图 2给出了引物UBC841的ISSR扩增产物在部分南方红豆杉种源植株样品中的分离情况。
|
|

|
图 2 引物UBC841的ISSR扩增产物在南方红豆杉部分植株中的分离 Figure 2 Segregation of ISSR bands amplified by UBC841 in some samples of T. wallichiana var. mairei |
ISSR为显性标记,同一引物扩增产物中电泳迁移率一致的条带被认为具有同源性。电泳图谱中的每一条带视为一个标记,并代表一个引物的结合位点。按照相同迁移位置上扩增条带的有无进行统计,有带的记为“1”,无带的记为“0”,仅记录清晰且长度在150~2 000 bp范围的扩增带,建立0/1数据矩阵,计算每个引物的多态性百分数和多态性信息含量(polymorphism information content, PIC)。应用POPGENE软件(Version 1.3.1)计算各种源的多态位点百分率(PPL)和Shannon信息多样性指数(I)、Nei's基因多样度指数(HE)(Nei, 1973)、物种水平基因多样性(HT)、种源内基因多样性(HS)、基因分化系数(GST)和基因流(Nm);用GenAlEx6对种源间和种源内分子遗传变异进行AMOVA分析(Peakall et al., 2006);用POPGENE软件计算种源间Nei’s(1972)遗传距离和Jaccard遗传相似系数,据此对15个种源进行UPGMA聚类分析,绘制种源的亲缘关系树状图。
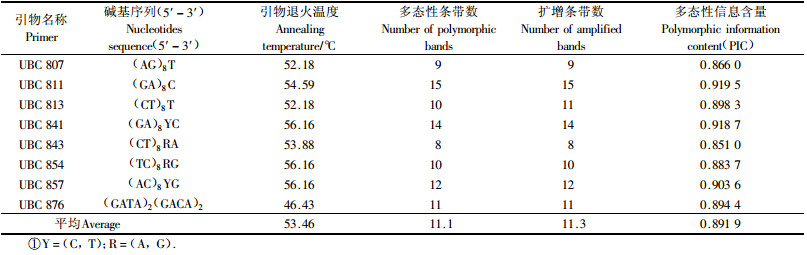
2 结果与分析 2.1 种源遗传多样性及地理变化利用8个ISSR引物对15个南方红豆杉种源共300个个体进行PCR扩增,共检测到90个位点,其中89个位点呈现多态性,多态性位点百分率(PPL)为98.89%(表 1),说明南方红豆杉物种水平上的遗传多样性丰富。每条引物扩增出的位点数在8~15个不等,平均为11.1个,条带片断大小介于150~2 000 bp间。8条引物的多态性信息含量变化在0.851 0~0.918 7,平均为0.891 9。
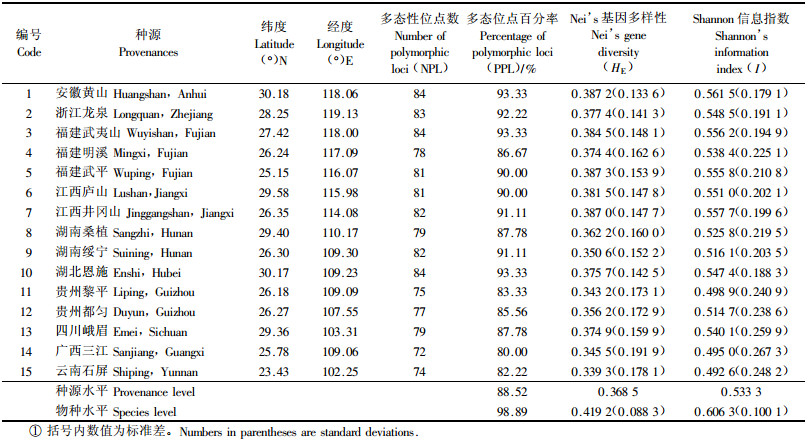
统计结果表明,15个南方红豆杉种源遗传多样性都处在较高水平。多态位点百分率(PPL)变化在80.00%~93.33%,平均为88.52%;Nei’s基因多样性变化在0.339 3~0.387 3,平均为0.368 5;Shannon信息指数(I)变化在0.492 6~0.561 5,平均0.533 3(表 2)。然而比较发现,不同南方红豆杉种源的遗传多样性还存在较大差异,安徽黄山,福建武夷山、武平,江西庐山和井冈山5个来自偏东和偏北部的种源遗传多样性高,HE和I分别大于0.38和0.55;而贵州黎平、广西三江和云南石屏3个来自偏西和偏南部的种源遗传多样性低,HE和I分别低于或接近于0.35和0.50;偏东和偏北部的5个种源较偏西和偏南部3个种源的HE和I的平均值提高了12.49%和12.29%。种源平均水平的各遗传多样性参数PPL为88.52%,HE为0.368 5,I为0.533 3;物种水平各遗传参数PPL为98.89%,HE为0.419 2,I为0.606 3。
|
|
从南方红豆杉不同种源Shannon信息指数(I)随产地经纬度变化的三维离散分布图(图 3)中可以清楚地看出,遗传多样性高的种源多分布在109°—120°E、26°—31°N范围内,包括安徽黄山、浙江龙泉等9个种源,其I平均值为0.550 7,较其他6个偏西和偏南种源的I均值(0.507 2)高出8.58%。构建了南方红豆杉种源I与产地经度(X1)和纬度(X2)的回归方程I=0.293 43-0.001 04X1X2+0.000 14X12+0.002 18X22,其决定系数R2值为0.816,达极显著水平。偏回归系数的F值检验结果表明,产地经度和纬度交互项及纬度平方项在回归方程中作用显著,意味着南方红豆杉种源遗传多样性是受其产地经度和纬度非线性共同影响。南方红豆杉自然分布区的偏东和偏北地区其天然林资源较多,种源遗传多样性较高,推测该区域可能是南方红豆杉的分布中心。偏南和偏西地区的南方红豆杉种源遗传多样性较低,也与现有天然林种群破坏严重有关(张宏意等, 2003)。

|
图 3 15个南方红豆杉试验种源Shannon信息指数(I)随产地经纬度的地理变化 Figure 3 Geographical change of Shannon's information index (I) for 15 tested provenances of T. wallichiana var. mairei with the latitude and longitude of the seed sources |
POPGENE分析表明南方红豆杉总的种源基因多样性为0.419 2,种源内基因多样性为0.368 5,种源间基因多样性为0.050 7,基因分化系数0.121 1,基因流很大,达3.627 7。
AMOVA分析结果(表 3)显示,南方红豆杉种源间的遗传变异较小,而种源内遗传变异显著(P < 0.001)。种源间遗传变异占总变异的8.75%,种源内变异占91.25%,与利用POPGENE软件分析获得的结果基本一致。
|
|

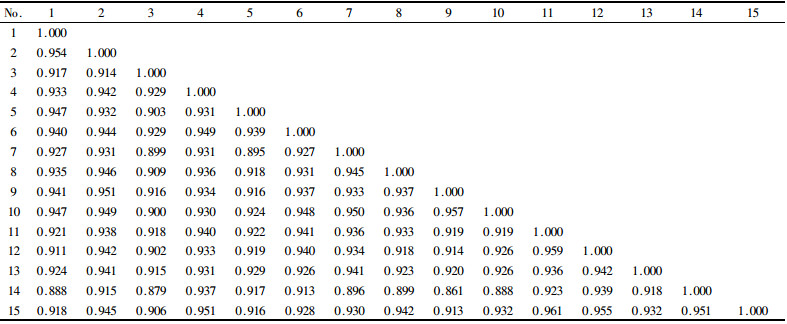
通过估算15个南方红豆杉种源间的遗传距离,并据此使用UPMGA聚类法得出树形图(图 4)。结果表明,15个南方红豆杉种源间遗传相似指数较大,介于0.861~0.959(表 4),意味着南方红豆杉种源间遗传差异不显著。从聚类图中可以看到,除福建武平和武夷山2个种源外,其他13个种源可明显地按地域聚为2组,一组包括安徽黄山、浙江龙泉、湖南绥宁、湖北恩施、江西井冈山和湖南桑植6个来自南方红豆杉自然分布区偏东或偏北的种源,其中安徽黄山与浙江龙泉,湖南绥宁与湖北恩施,江西井冈山与湖南桑植种源的遗传一致性较大,分子相似指数分别为0.954、0.957和0.945;另一组包括贵州黎平、云南石屏、都匀、江西庐山、福建明溪、广西三江和四川峨眉等7个来自偏南或偏西的种源,其中贵州黎平、都匀和云南石屏种源间的遗传距离最近,分子相似指数皆大于0.955。福建武平和武夷山2个原产地种群都较小的种源虽然都维持了较高的遗传多样性,但与上述2组种源间的遗传距离都相对较远,另单独聚成2组。

|
图 4 15个南方红豆杉试验种源Nei’s(1972)遗传距离的UPGMA聚类图 Figure 4 UPGMA dendrogram based on Nei's(1972) genetic distances between 15 tested provenances of T. wallichiana var. mairei |
|
|
Hamrick等(1990)总结了80种裸子植物遗传多样性的研究结果,发现裸子植物的遗传变异性较高,平均而言,多态位点比率为0.709;对一些珍稀濒危物种而言,遗传多样性水平可能要低一些,如银杉(Cathaya argyrophylla)3个居群的多态位点仅为0.200~0.280(葛颂等, 1997)。北美脂松(Pinus resinosa)的遗传多样性则更低,多态位点比率仅为0.111(Allendorf et al., 1982)。本文对南方红豆杉15个种源的遗传多样性研究结果表明,种源平均多态性位点为0.885,说明南方红豆杉种源的遗传多样性非常丰富,总的基因多样性为0.419 2,这不仅显著地高于Nybom(2004)所统计的多种植物种群水平的遗传多样性(基于RAPD和ISSR等显性标记)平均值(HE=0.22),而且也明显地高于具有与南方红豆杉相似分布区的我国珍稀濒危树种三尖杉(Cephalotaxus fortunei)(HE=0.337 7)(李因刚等,2008)等。试验的15个种源皆维持了较高水平的遗传多样性,平均Nei’s基因多样性(HE)为0.368 5,福建武平和武夷山2个东部种源虽然种群较小,但其Nei’s基因多样性(HE)仍分别高达0.387 3和0.384 5,这说明南方红豆杉濒危的原因不是由于其遗传多样性的丧失,而是与其生境要求严格、天然更新能力弱以及对其过度采伐利用等有关。研究组对红豆树(Ormosia hosiei)自然保留种群遗传多样性的研究也获得了类似的结果(赵颖等, 2008)。南方红豆杉维持较高水平的遗传多样性,应与其呈群状广域性分布、风媒异花授粉、寿命长而世代重叠等特性有关。
南方红豆杉属我国一级珍稀濒危保护树种,虽然种源间的生长和形态差异显著(焦月玲等, 2007),但在DNA水平上种源间遗传距离较近,基因分化系数为0.121 1,有8.75%的遗传变异存在于种源间,而种源内的变异占总变异的91.25%。与三尖杉(李因刚等,2008)等我国亚热带地区的其他珍稀濒危树种比较,南方红豆杉种源间遗传分化较小,这不仅与各种源维持较高水平的遗传多样性有关,而且与其传粉和种子传播能力强及现有保留种群片断化时间短等有关。南方红豆杉现有保留种群一般都较大,多呈集群分布,与三尖杉种群小且呈星散分布等不同(李因刚等,2008),这些保留种群皆是在原有大量天然林资源过度采伐利用后保留下来的古树群,树龄皆在百年甚至千年以上。南方红豆杉为风媒异花授粉树种,传粉能力强,其种子具有红色带甜味的肉质假种皮,可借助鸟及鼠类的吞食而得以远距离传播,原有种群间的基因交流频繁,加之现有保留种群片断化的时间较短,未发生严重的遗传分化。
南方红豆杉不同种源虽然遗传多样性都较高,遗传距离较近,但仍存在明显的地理变异模式。基于UPMGA聚类结果,除个别较小种群的种源外,南方红豆杉种源可大致按地域聚为2组。偏东和偏北部的种源聚为一组,其遗传多样性普遍较高;偏南和偏西部的种源聚为另一组,其遗传多样性普遍较低。从南方红豆杉野生资源分布情况来看,中东部地区的野生资源量多,分布广,种群大,而偏南和偏西地区的野生资源量少,分布较零星,种群较小,这应是偏东和偏北部种源遗传多样性高而偏南和偏西部种源遗传多样性低的重要原因之一。张宏意等(2003)对于广东、湖南和江西较小分布区12个南方红豆杉自然种群遗传多样性的研究结果也证实了这一点,来自偏南地区也即来自粤北的种群遗传多样性较低。南方红豆杉自然分布区之中东部地区的种源遗传多样性较高,应优先加强这些地区南方红豆杉自然资源,尤其是较大自然保留种群的保护和优良种质的开发利用。
葛颂, 王海群, 张灿明, 等. 1997. 八面山银杉林的遗传多样性和群体分化. 植物学报, 39(3): 266-271. |
焦月玲, 周志春, 余能健, 等. 2007. 南方红豆杉苗木性状种源分化和育苗环境影响. 林业科学研究, 20(3): 363-369. DOI:10.3321/j.issn:1001-1498.2007.03.012 |
李因刚, 周志春, 金国庆. 2008. 三尖杉种源遗传多样性. 林业科学, 44(2): 64-69. DOI:10.3321/j.issn:1001-7488.2008.02.010 |
张宏意, 陈月琴, 廖文波. 2003. 南方红豆杉不同居群遗传多样性的RAPD研究. 西北植物学报, 23(11): 1994-1997. DOI:10.3321/j.issn:1000-4025.2003.11.027 |
赵颖, 何云芳, 周志春, 等. 2008. 浙闽5个红豆树自然保留种群的遗传多样性. 生态学杂志, 27(8): 1279-1283. |
周其兴, 葛颂, 顾志建, 等. 1998. 中国红豆杉属及其近缘植物的遗传变异和亲缘关系分析. 植物分类学报, 36(4): 323-332. |
Allendorf F W, Knudsen K L, Balk G M. 1982. Frequencies of null alleles at enzyme loci in natural populations of ponderosa and red pine. Genetics, 100: 497-504. |
Doyle J J, Doyle J L. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf material. Phytochem Bull, 19: 11-15. |
Fu L, Mill R R. 1999. Taxaceae. Flora of China, 4: 89-96. |
Hamrick J L, Godt M J W. 1990. Allozyme diversity in plant species//Brown A D H, Clegg M T, Kahlar A C, et al.Plant population genetics, breeding, and genetic resources. Sunderland Mass: Sinauer, 43-63.
|
Lewandowski A, Burczyk J, Mejnartowicz L. 1995. Genetic structure of English yew (Taxus baccata L.) in the Wierzchlas Reserve: implications for genetic conservation. Forest Ecology and Management, 73: 221-227. DOI:10.1016/0378-1127(94)03477-E |
Li X L, Yu X M, Guo W L. 2006. Genomic diversity within Taxus cuspidata var. nana revealed by random amplified polymorphic DNA markers. Russian Journal of Plant Physiology, 53: 771-776. |
Nei M. 1972. Genetic distance between populations. Amer Naturalist, 106: 283-292. DOI:10.1086/282771 |
Nei M. 1973. Analysis of gene diversity in subdivided populations. Proc Natl Acad Sci USA, 70: 3321-3323. DOI:10.1073/pnas.70.12.3321 |
Nybom H. 2004. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Molecular Ecology, 13: 1143-1155. DOI:10.1111/mec.2004.13.issue-5 |
Peakall R, Smouse P E. 2006. GenAlEx 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes, 6: 288-295. DOI:10.1111/men.2006.6.issue-1 |
Senneville S, Beaulieu J, Daoust G, et al. 2001. Evidence for low genetic diversity and metapopulation structure in Canada yew (Taxus Canadensis): considerations for conservation. Can J For Res, 31: 110-116. |
Shah A, Li D Z, Gao L M, et al. 2007. Genetic diversity within and among populations of the endangered species Taxus fauna (Taxaceae) from Pakistan and implications for its conservation. Biochemical Systematics and Ecology, 7: 1-11. |