 2009, Vol. 45
2009, Vol. 45文章信息
- 张智婷, 宋新章, 肖文发, 高宝嘉, 张慧玲, 胡雁林, 国立红.
- Zhang Zhiting, Song Xinzhang, Xiao Wenfa, Gao Baojia, Zhang Huiling, Hu Yanlin, Guo Lihong
- 长白山杨桦次生林采伐林隙种子库特征
- The Seed Bank in Logging Gaps in Populus davidiana-Betula platyphylla Secondary Forests in Changbai Mountain
- 林业科学, 2009, 45(1): 21-26.
- Scientia Silvae Sinicae, 2009, 45(1): 21-26.
-
文章历史
- 收稿日期:2007-10-15
-
作者相关文章



2. 浙江林学院 杭州 311300;
3. 中国林业科学研究院森林生态环境与保护研究所 北京 100091;
4. 吉林省白河林业局 安图 133613
2. Zhejiang Forestry University Hangzhou 311300;
3. Institute of Forest Ecology, Environment and Protection, CAF Beijing 100091;
4. Baihe Forestry Bureau Antu 133613
土壤种子库作为一个潜在的群落体系,在森林生态系统生物多样性维持中具有重要作用(Grubb, 1996;Huston, 1994)。土壤种子库中种子的多度和多样性在很大程度上决定着未来幼苗、幼树的多度和多样性,进而决定着森林群落的物种丰富度和多样性特征(臧润国等,2004)。了解林隙内种子库的数量和分布,有助于理解林隙更新动态在群落更新与物种多样性动态维持中的作用。在大多数的林隙动态研究中,特别是热带林的研究中,由于研究难度较大,种子往往被忽略不计(Brokaw, 1985;Zang et al., 2002; Hubbel et al., 1999)。目前,国内外有关林隙与种子库动态的研究还不多(Zang et al., 2008; Connell, 1989; Denslow et al., 1990),研究深度也不够。但许多植物种群学家和森林动态学家都充分认识到了林隙内的种子库是森林植物种群动态和森林更新的一个重要环节,是森林生物多样性维持机制的一个重要方面,近几年来有关这方面的研究逐渐增加(臧润国等, 2004; Grubb, 1996)。天然次生林已成为我国森林资源的主体(朱教君, 2002),鉴于次生林在我国森林资源和生态环境建设中的重要地位和作用,进行次生林林隙更新动态方面的研究,探讨次生林演替的内在生态学机理,对于实现次生林的可持续经营具有重要意义。
1 研究区概况研究在吉林省白河林业局天然杨桦(Populus davidiana-Betula platyphylla)次生林经营采伐迹地进行(42°05′—42°28′ N,128°02′—128°64′ E)。该区属温带季风影响下的大陆性季风气候,年平均气温7.3~4.9 ℃,年降水量600~900 mm,全年日照时数为2 271~2 503 h,全年无霜期109~141 d。海拔700 m,坡度1°~5°,为比较平缓的平地与较浅的沟谷相交错的地形。土壤为火山灰形成的典型暗棕壤,土层厚100 cm以上。林龄50 a,属中龄林,分别于2002年和1996年进行了人工择伐,择伐后形成了大小不一的林隙,林分密度556株·hm-2,林分蓄积量为110 m3·hm-2,平均直径16 cm,平均树高15 m,郁闭度0.6。主要树种有山杨(Populus davidiana)、白桦(Betula platyphylla)、色木槭(Acer mono)、胡桃楸(Juglans mandshurica)、山荆(Malus baccata)、椴木(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、山槐(Maackia amurensis)、蒙古栎(Quercus mongolica)和春榆(Ulmus japonica)等, 主要灌木有忍冬(Lonicera ruprechtiana)、暴马丁香(Syringaamurensis)、卫矛(Euonymus alatus)、榛子(Corylus heterophylla)等。主要草本植物为莎草(Cyperus rotundus),盖度30%,分布均匀。
2 研究方法 2.1 调查方法在大面积踏查的基础上,将采伐林隙的面积分为5级:< 50,50~100,100~150,150~200和200~250 m2。选择2002年采伐形成的5个有代表性的大小级别的椭圆形林隙各1个,林隙面积分别为42,85,106,188和236 m2;1996年的采伐林隙较少,加上天气原因,只调查了2个林隙,面积分别为66和138 m\+2。调查林隙边缘木的胸径、树高等因子。在每个扩展林隙内设9个2 m×2 m的样方,第1个样方设在林隙中心,其余8个分别沿长短轴方向设在距离扩展林隙边缘的一半处和扩展林隙边缘处。在每个小样方内进行土壤种子库调查。同时,在距林隙边缘10 m处的非林隙林分中相应设1个10 m×10 m的对照样地,对照样地郁闭度为0.8,在每个对照样地内沿对角线方向机械设9个2 m×2 m的样方进行种子库调查。调查于2006年5月16—20日进行。该时期长白山林区土壤刚刚解冻,种子尚未萌发,基本不受地表杂草的影响,比较适合种子库的取样。在各样方内用1个0.5 m×0.5 m的木质采样框固定采样面积,分枯枝落叶层和腐殖质层(地表 0~10 cm)采样,收集框内的全部地表凋落物和地表 0~10 cm的土样分别装入布袋,做好标记,带回室内进行土壤筛选。对每份样品进行室内逐点逐层过筛、淘洗。首先将每份土样依次过3与2 mm筛,然后挑选出所有种子;最后进行淘洗,待反复冲洗干净后剔除杂物,拣出各种植物种子进行统计。对照种子标本进行种子鉴定。根据研究需要,本研究只统计木本植物的种子,不进行生活力的测定。用种子库的丰富度和个体密度来表示土壤种子库的储量特征。为便于表述,文中分别用L,O和L+O层来表示枯枝落叶层、腐殖质层和两层共有的种子库。
另外,本研究还进行了种子库同更新幼苗相似性的比较。2005年已在上述每个2 m×2 m样方内进行了更新幼苗调查,更新幼苗调查方法详见宋新章等(2008)。
2.2 多样性指数计算采用5类目前普遍应用的多样性指数来测定林隙内土壤种子库的物种多样性(臧润国等, 1999;2002; 刘金福等, 2003; 吴承祯等, 1996; 洪伟等, 1999)。物种丰富度指数(R)计算公式为R=(S-1)/lnN,式中:S为物种总数即丰富度,N为样方内观察到的个体总数;物种多样性Shannon-Wiener指数(H)计算公式为
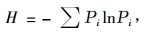
|
式中:Ρi是第i物种的个体数占总个体数N的比例;群落均匀度Pielou指数(Js)计算公式为
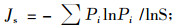
|
生态优势度Simpson指数(λ)计算公式为
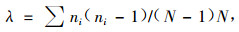
|
式中:ni是第i物种的个体数;相似性系数Jaccard指数(Z)计算公式为Cj=j/(a+b-j),式中:j为2个群落或样地共有种数,a和b分别为2个群落或样地各自的物种数;相异性系数(CD)计算公式为CD=1-2j/(a+b)。
3 结果与分析 3.1 土壤种子库的丰富度和个体密度由表 1可以看出,杨桦次生林各大小级林隙内土壤种子库的丰富度存在较大差异。3龄林隙中,50~200 m2林隙的L,O和L+O层的丰富度均较高,尤其是50~100 m2最高,L+O层达27.1种·m-2,< 50 m2林隙内各层种子库的丰富度均最低,L+O层的丰富度仅19.1种·m-2,为50~100 m2林隙的70.5%。3龄林隙的非林隙林分内种子库的丰富度除稍高于 < 50 m2林隙外,均低于其他大小级林隙。9龄林隙中也是50~100 m2林隙L+O层种子库的丰富度最大,达23.1种·m-2,但低于3龄林隙。9龄林隙的非林隙林分内种子库的丰富度不但低于9龄林隙,也远低于3龄林隙及其非林隙林分。
|
|
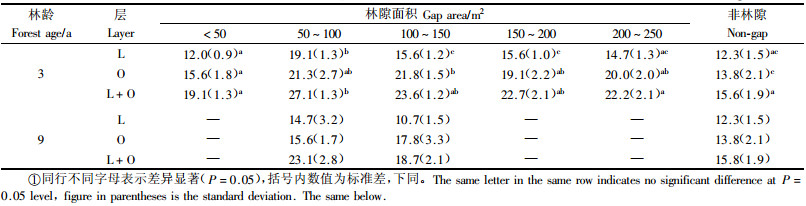
由表 2可以看出,各大小级林隙内种子库的个体密度也存在较大差异。3龄林隙50~100 m2林隙内L,O和L+O层的个体密度均显著大于其他大小级林隙,L+O层为511.1个·m-2。150~200 m2林隙内L+O层的个体密度最低,为181.3个·m-2,仅为50~100 m2林隙的35.5%。非林隙林分种子库L层的个体密度均高于林隙,O和L+O层的个体密度除低于50~100 m2林隙外,均高于其他大小级林隙。
|
|
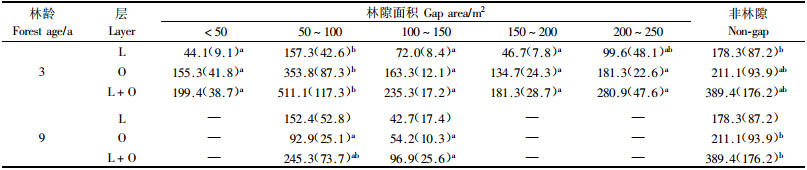
9龄林隙中,50~100 m2林隙内种子库各层的个体密度最大,L+O层为245.3个·m-2,100~150 m2林隙内种子库的个体密度最小,L+O层为96. 9个·m-2,仅为50~100 m2林隙的39.5%。与9龄林隙相邻的非林隙林分种子库各层的个体密度则远远大于9龄各大小林隙。
从表 1和2可以看出,不论是3龄还是9龄林隙,50~100 m2林隙内种子库各层的丰富度和个体密度均最高,表明该大小的林隙较有利于种子储存,具体原因尚不清楚。9龄林隙内种子库各层的丰富度和个体密度均低于相应大小的3龄林隙。表明随着林隙形成时间的延长,林隙种子库的种子储量逐渐减少。这可能与生境改变,刺激了种子大量萌发有关。
由图 1可见,杨桦次生林3和9龄林隙内不同部位间L,O和L+O层种子库的丰富度和个体密度变化较大,并无明显的规律性。3龄林隙内中南部L+O层的丰富度最大,达28种·m-2,林隙东缘和中西部L+O层的丰富度最低,为20种·m-2,为中南部的71.4%;林隙西缘L+O层的个体密度最高,达483个·m-2,林隙中南部L+O层的个体密度最低,仅147.3个·m-2,为西缘的30.4%。9龄林隙北部L+O层的丰富度最大,达32种·m-2,林隙南缘L+O层的丰富度最低,为10种·m-2,为中北部的31.2%;林隙东缘L+O层的个体密度最高,达364个·m-2,林隙南缘L+O层的个体密度最低,仅40个·m-2,为东缘的11%。可见,9龄林隙东缘种子库的丰富度和个体密度均较高,而南缘种子库的丰富度和个体密度均最低。3龄林隙内各部位O层的丰富度和个体密度都大于L层,而9龄林隙内各部位L层的个体密度均高于O层,这是由于9龄林隙内的凋落物量较大,种子掉落到地表时受凋落物层的阻力也加大,从而被更多地滞留在了凋落物层。总体来看,3龄各部位种子库L+O层的丰富度和个体密度的变化趋势与O层种子库基本一致,说明3龄林隙种子库的数量和组成主要由O层决定, 而9龄林隙种子库L+O层的丰富度和个体密度的变化趋势与L层种子库基本一致,说明9龄林隙种子库的数量和组成主要由L层决定。

|
图 1 林隙内土壤种子库的空间差异 Figure 1 Spacial heterogeneity of soil seed bank in gaps CC:林隙中心Gap center;NN:扩展林隙北部林缘处North edge of expanded canopy;CN:林隙中心至林隙北部林缘距离的一半处Center of gap center to north edge;CS:林隙中心至南部林缘距离的一半处Center of gap center to south edge;SS:扩展林隙南部林缘处South edge of expanded canopy;EE:扩展林隙东部林缘处East edge of expanded canopy;CE:林隙中心至东部林缘距离的一半处Center of gap center to east edge;CW:林隙中心至西部林缘距离的一半处Center of gap center to west edge;WW:扩展林隙西部林缘处West edge of expanded canopy;NG:非林隙Non-gap. |
由表 3可以看出,杨桦次生林3龄不同大小林隙L+O层种子库中主要树种的个体密度差异巨大。50 m2以下的林隙种子库中,山槐、色木槭和蒙古栎的密度最大,这3个树种的种子数量占种子总量的比例达72%。50~100 m2林隙种子库中,紫椴、蒙古栎和色木槭的密度最大,这3个树种的种子数量占种子总量的79.4%以上。100~150 m2林隙种子库中,紫椴(Tilia amurensis)、色木槭、山槐和蒙古栎的密度最大,这4个树种的种子数量占种子总量的85.9%。150~200 m2林隙种子库中,山槐和色木槭的密度最大,这2个树种的种子数量占到种子总量的75.7%。200~250m2林隙种子库中,水曲柳、色木槭和山槐的密度最大,这3个树种的种子数量占种子总量的87.7%以上。对比各大小林隙可知,色木槭和山槐在各大小级林隙内种子库中的密度均较高,且都占有重要地位,蒙古栎、紫椴和水曲柳只在个别林隙种子库中占主要地位,其余树种的密度则在各大小级林隙内无太大变化,甚至只存在于个别林隙的种子库中,如红松(Pinus koraiensis)、黄檗(Phellodendron amurense)等。
|
|

总体来看,林隙L+O层种子库中紫椴、色木槭、山槐和蒙古栎的个体密度最大,这4个树种的种子数量占种子总量的79%以上;非林隙林分的土壤种子库中,紫椴、水曲柳、色木槭和山槐的密度最大,这4个树种的种子数量占到种子总量的90.4%,林隙内外的对比显示,除蒙古栎和糠椴外,各主要树种在非林隙林分种子库中的密度均要大于林隙内。
3.3 3龄林隙土壤种子库多样性由表 4可见,杨桦次生林3龄不同大小林隙内L+O层种子库的多样性存在较大差异。50 m2以下林隙种子库的Shannon-Wiener指数H最高,可达1.83,表明该大小林隙种子库的物种多样性和综合多样性最大。150~200 m2林隙种子库的Margalef指数R最高,为2.50,表明该大小林隙种子库的物种丰富度最高。100~150 m2林隙种子库的Pielou指数Js最大,为0.79,表明该大小林隙种子库的物种均匀度较好。200~250 m2以上的林隙种子库的Simpson指数λ最大,达0.29,表明该大小林隙种子库的物种优势度较高。
|
|

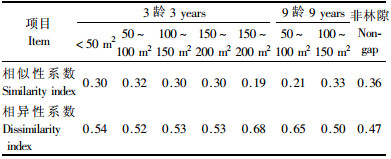
用相似性系数Jaccard指数(Cj)初步测度了同一林型不同大小、不同年龄林隙L+O层种子库的相似性与相异性,以探讨林隙大小和形成时间长短对种子库物种组成的影响。由表 5可以看出,杨桦次生林3龄各大小林隙土壤种子库的物种组成较相似,相似性系数均达0.5以上,且表现出林隙面积差异愈小,相似性系数愈大的趋势。其中50~100 m2林隙内的种子库同 < 50 m2和150~200 m2林隙内的种子库物种组成相似性系数最大,为0.69,表明此类大小林隙内种子库的组成较相似;< 50 m2同150~200 m2林隙内的种子库物种组成相似性系数最低,为0.5,表明此类大小林隙内种子库的物种组成差异较大。9龄林隙内土壤种子库的物种组成同3龄林隙中面积较小林隙(100 m2以下)的种子库差异较大,相似性系数均在0.5以下,而同面积较大林隙(100 m2以上)种子库的差异较小,相似性系数均在0.5以上。综合来看,在杨桦次生林采伐林隙形成初期,林隙形成时间较之林隙大小对林隙内土壤种子库物种组成的影响更大。
|
|

由表 6可以看出,杨桦次生林3龄林隙L+O层种子库内,200 m2以下各大小林隙种子库的物种组成同更新幼苗的相似性系数均较低,大多为0.3,而相异性系数均较高,一般在0.52左右。150~200 m2林隙内种子库物种组成同更新幼苗的相似性更低,仅为0.19,相异性系数则相对较高,为0.68。9龄林隙内种子库组成同幼苗的相似性也不高。非林隙林分种子库的物种组成同幼苗的相似性系数高于林隙,相异性系数则低于林隙。表明在采伐林隙形成初期,采伐干扰对种子库的组成、幼苗的萌生和存活有着重要影响,而大强度干扰产生的影响也更大。
|
|
本研究分析长白山区杨桦次生林不同大小采伐林隙的土壤种子库特征。结果表明:非林隙林分中种子库储量远高于林隙内,但多样性比林隙低。不同大小林隙的种子库储量差异显著,其中3龄林隙50~100 m2林隙内种子库储量最大。随林隙形成时间延长,林隙内种子库储量减少,但多样性增加。这可能与林隙形成后,原有的种子逐渐萌发,而新落入的种子数量逐渐减少有关。林隙内不同部位间的种子库储量变化较大,种子的丰富度和个体密度变化不一致,即种类多的种子库中种子的数量不一定多,而数量大的种子库中的种子种类不一定多,这可能同林隙周边母树的分布有关。种子库中水曲柳、山槐和榛子的密度最大,这3个树种的种子数量占种子总量的74.7%以上。值得注意的是,本研究虽然是在次生杨桦林内进行,但却没有发现白桦和山杨的种子,这可能同取样的季节有关。事实上,对林隙内更新幼苗的调查也表明白桦和山杨的幼苗几乎不存在(宋新章等, 2008)。
在林隙形成初期,种子库组成及数量主要由腐殖质层决定,这可能是因为此时凋落物少,对种子落入土壤的阻力较小,种子能顺利地进入腐殖质层;随着时间推移,林隙内累积的凋落物数量逐渐增多,此时凋落物层就成为种子落入土壤的一大障碍,使得种子更多地滞留在了凋落物层,从而使凋落物层种子库对整个种子库的作用越来越大。王辉等(2004)对子午岭主要森林类型土壤种子库的研究也表明,针叶树的种子多集中在枯枝落叶层和0~2 cm土层,向下种子数量递减,可能与针叶林枯落层普遍较厚有关。肖智术等(2001)的研究也发现土壤种子库各组成主要分布在土壤表面的枯枝落叶物层和浅层土壤中。这表明森林土壤种子库中种子的垂直分布具有一定的规律性,即多数种子滞留在枯枝落叶层和浅层土壤中,深层土壤中的种子含量较少。
林隙内种子库多样性要高于非林隙,但种子的生态优势度低于非林隙,可能是由于采伐干扰造成的小生境异质性促进了种子多样性增加。宋新章等(2008)对该杨桦次生林样地幼苗更新状况的调查研究表明,采伐林隙内更新幼苗的种数和个体密度均高于非林隙林分。张春雨等(2006)对长白山阔叶红松林林隙与林下更新幼苗状况的研究也表明,同林下相比,林隙中幼苗层树种多样性增大而群落优势度减小。可见,土壤种子库的多样性对幼苗更新的多样性有一定的影响。
同非林隙相比,林隙内土壤种子库组成同更新幼苗的相似性较低。黄忠良等(1996)对鼎湖山不同演替阶段森林土壤种子库的研究结果表明地上植物种类与土壤种子库组成的相似性不论在演替早期,还是在中后期都不明显。唐勇等(1999)发现西双版纳热带幼龄次生林土壤种子库与地上植被的共有种类较多,但是随着林龄增加,土壤种子库与地上植被共有种类的储量明显下降。多数研究亦表明,种子库组成同地上植被的相似性不明显(Hall et al., 1980; Balum, 1993)。
由于条件限制,本研究只在生长季开始前进行种子库的取样工作,事实上秋天树木结实下落后,有相当一部分种子被林内动物啃食掉或储藏起来,而且某些动物对种子的习惯性偏好和选择性也会使某些树种的种子损失严重,此时的种子库已不能全面反映真实的种子库状况。而秋冬季受大雪阻碍,亦不利于种子库的调查。另外,本研究只对种子的数量和种类进行了统计分析,而没有对种子的生活力进行鉴定,这也限制了对林隙内种子库状况及其对幼苗更新影响的全面认识。以后应开展林隙内种子雨及种子生活力的研究。
洪伟, 林成来, 吴承祯, 等. 1999. 福建建溪流域常绿阔叶防护林物种多样性研究. 生物多样性, 8(3): 208-213. DOI:10.3321/j.issn:1005-0094.1999.03.008 |
黄忠良, 孔国辉, 魏平, 等. 1996. 南亚热带森林不同演替阶段土壤种子库的初步研究. 热带亚热带植物学报, 4: 42-49. |
刘金福, 于玲, 洪伟, 等. 2003. 格氏栲林林窗物种多样性动态规律的研究. 林业科学, 39(6): 159-164. DOI:10.3321/j.issn:1001-7488.2003.06.027 |
宋新章, 张智婷, 肖文发, 等. 2008. 长白山杨桦次生林采伐林隙幼苗更新动态. 林业科学, 44(3): 13-20. |
唐勇, 曹敏, 张建侯, 等. 1999. 西双版纳热带森林土壤种子库与地上植被的关系. 应用生态学报, 10(3): 279-282. DOI:10.3321/j.issn:1001-9332.1999.03.006 |
王辉, 任继周. 2004. 子午岭主要森林类型土壤种子库研究. 干旱区资源与环境, 18(3): 130-136. DOI:10.3969/j.issn.1003-7578.2004.03.026 |
吴承祯, 洪伟, 陈辉, 等. 1996. 万木林中亚热带常绿阔叶林物种多样性研究. 福建林学院学报, 16(1): 343-347. |
肖治术, 王玉山, 张知彬. 2001. 都江堰地区三种壳斗科植物的种子库及其影响因素研究. 生物多样性, 9: 373-381. DOI:10.3321/j.issn:1005-0094.2001.04.008 |
臧润国, 安树青, 陶建平, 等. 2004. 海南岛热带林生物多样性维持机制. 北京: 科学出版社.
|
臧润国, 蒋有绪, 余世孝. 2002. 海南霸王岭热带山地雨林森林循环与树种多样性动态. 生态学报, 22(1): 24-32. DOI:10.3321/j.issn:1000-0933.2002.01.004 |
臧润国, 徐化成, 高文韬. 1999. 红松阔叶林主要树种对林隙大小及其发育阶段更新反应规律的研究. 林业科学, 35(3): 90-98. |
张春雨, 赵秀海, 郑景明. 2006. 长白山阔叶红松林林隙与林下土壤性质对比研究. 林业科学研究, 19(3): 347-352. DOI:10.3321/j.issn:1001-1498.2006.03.016 |
朱教君. 2002. 次生林经营基础研究进展. 应用生态学报, 13(12): 1689-1694. DOI:10.3321/j.issn:1001-9332.2002.12.040 |
Balun L A. 1993. Comparative soil seed bank study of four vegetation types at Bulolo, Papua New Guinea. Science-in-New-Guinea, 19(2): 87-96. |
Brokaw N V L. 1985. Gap-phase regeneration in a tropical forest. Ecology, 66: 682-687. DOI:10.2307/1940529 |
Connell J H. 1989. Some processes affecting the species composition in forest gaps. Ecology, 70: 560-562. DOI:10.2307/1940205 |
Denslow J S, Gomez Diaz A E. 1990. Seed rain to tree-fall gaps in a Neotropical rain forest. Canada Journal of Forest Research, 20: 642-648. DOI:10.1139/x90-086 |
Grubb P J. 1996. Rainforest dynamics: the need for new paradigm//Edwards D S, Booth W E, Choy S C. Tropical rainforest research-current issues. Boston: Kluwer Academic Publishers, 215-234.
|
Hall J B, Swaine M D. 1980. Seed stocks on Ghanaian forest soil. Biotropica, 12: 256-263. DOI:10.2307/2387695 |
Hubbel S P, Foster R B, O Brien S T, et al. 1999. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science, 283: 554-557. DOI:10.1126/science.283.5401.554 |
Huston M A. 1994. Biological diversity-The coexistence of species on changing landscapes. New York: Cambridge University Press.
|
Zang R G, Ding Y, Zhang W Y. 2008. Seed dynamics in relation to gaps in a tropical montane rainforest of Hainan island, South China: (Ⅱ) seed bank. Journal of Integrative Plant Biology, 50(5): 513-521. DOI:10.1111/jipb.2008.50.issue-5 |
Zang R G, Wang B S. 2002. Study on canopy disturbance regime and mechanism of tree species diversity maintenance in the lower subtropical evergreen broad-leaved forest, South China. Plant Biosystems, 136(2): 241-250. DOI:10.1080/11263500212331351149 |