 2008, Vol. 44
2008, Vol. 44文章信息
- 张淑勇, 周泽福, 张光灿, 夏江宝.
- Zhang Shuyong, Zhou Zefu, Zhang Guangcan, Xia Jiangbao
- 半干旱黄土丘陵区4种天然次生灌木光合生理和水分利用特征
- Characteristics of Photosynthesis, Respiration and Water Utility of Four Tree Species of Natural Secondary Shrubs in a Semi-arid Loess Hilly-gully Region
- 林业科学, 2008, 44(12): 140-146.
- Scientia Silvae Sinicae, 2008, 44(12): 140-146.
-
文章历史
- 收稿日期:2007-12-27
-
作者相关文章



2. 山东农业大学林学院 泰安 271018;
3. 滨州学院黄河三角洲生态环境研究中心 滨州 256603
2. Forestry College, Shandong Agricultural University Tai'an 271018;
3. Research Center for Eco-Environmental Sciences of Yellow River Delta, Binzhou University Binzhou 256603
在黄土高原地区,干旱缺水是制约当地经济发展的突出生态特征,也是制约植被恢复和农林业生产重要因素(王克勤等,1998;程积民等,2001;李世东等,2002;安韶山等,2006)。黄土高原植被的恢复过程关键在于树种的选择,以灌木为主,营造灌、草、乔相结合的水土保持林,对加快绿化荒山荒沟速度、保持水土具有重要的现实意义(邹厚远等,1995;程积民等,2001;李世东等,2002;万雪琴等,2005)。但是,从黄土高原地区人工植被建设与退耕还林的现状看,很多地区仍然存在树种单一、树种选择与立地配置不合理的问题。主要原因之一是人们对当地一些可以开发利用的优良树种资源认识不够,缺乏相关植物种类的水分关系研究(程积民等,2001;胡良军等,2002;李世东等,2002;万雪琴等,2005;孙志蓉等,2006)。
黄刺玫(Rosa xanthina)、柠条(Caragana korshinskii)、山桃(Prunus davidiana)及山杏(Prunus sibirica)是半干旱黄土丘陵区天然次生林中广泛散生分布的灌木树种,它们具有一定的观赏、药用价值和良好的生态功能,在退耕还林与植被恢复工程中,具有较大的开发潜力。国内对柠条的生物学特性、生理生化特征以及抗旱适应性等方面均已进行了大量研究(唐海萍等,2001;牛西午等,2003;许炳成等,2004;安韶山等, 2006, 孙志蓉等,2006),有关黄刺玫、山桃及山杏的研究,多集中于栽培技术、解剖结构、林地水分动态及耗水性能方面(王孟本等,1999;汪天等,2000;王淼等,2001;程积民等,2003),但有关光合生理生态等方面的比较相对较少。本研究系统地比较4种灌木树种在半干旱黄土丘陵区的光合、蒸腾及水分利用等生理参数特征,可为该区在人工植被建设中的科学合理配置提供科学依据。
1 试验地概况研究区位于山西省中阳县的车鸣峪林场,属黄河流域典型的半干旱黄土丘陵区,111°04′—111°21′ E、37°03′—37°20′ N。该区属暖温带亚干旱区,干燥具明显的大陆性季风气候,多年平均降水量为500~550 mm,降雨集中在7—9月份(占全年降水量的70%以上),平均蒸发量为1 019.7 mm,平均气温6 ℃左右,极端最高气温35.6 ℃(1994年6月16日),极端最低气温-24.3 ℃(1980年1月30日),无霜期125~150 d,≥10 ℃年积温为2 500~3 000 ℃。土壤以褐土居多,其次是棕壤,发育差、水土流失严重。属于森林草原灌丛植被区,现有树种少,林地大部分是疏林地,林分稳定性差。
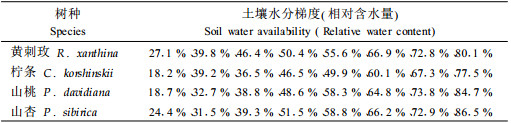
2 材料与方法 2.1 试验材料3年生天然次生灌木黄刺玫、柠条、山桃及山杏,4种苗木为天然次生林中筛选,苗木在全光照下生长,2007年3月从天然次生林中各选取6株(6个重复)进行苗木盆栽,盆内土壤为褐土,盆钵直径40 cm,深50 cm。8月中旬进行试验观测。用环刀法测得盆栽实验土壤的田间持水量和土壤密度(表 1)。
|
|

1) 水分处理 采用田间环境条件下人为给水和自然耗水相结合的方法,获取盆栽土壤不同的水分梯度,选取生长较一致健壮的盆栽苗各3株(每盆1株),试验观测2 d前浇水,使土壤水分饱和,利用美国产MI2X型土壤水分探头监测土壤体积含水量(voluminal water content,VWC;%)的变化。2 d后获得初期水分梯度,进行第1次生理参数的测定。以后通过自然耗水,依据D-最优性法则(Kiefer et al., 1959),每隔1~2 d获取1个水分梯度。按照RWC在表 2中水分点进行生理参数光响应过程的测定。
|
|
2) 光响应过程测定 选取盆栽苗木3株,从试验植株中部分别选3片生长健壮的成熟叶片,8月中旬用Li-6400便携式光合仪测定不同水分梯度下生理参数的光响应过程,为尽量减少外界光照波动所造成的影响,测定时间选择在完全晴朗天气时的8:00—10:00左右时进行,每个叶片重复3次,取平均值。测定时,使用大气CO2浓度(365±5) μmol·mol-1,叶室温度由Li-6400温度控制器控制在(25±0.5)℃。利用人工光源将光合有效辐射强度设置2 000,1 700,1 400,1 100,900,700,500,300,150,100,50,20 μmol·m-2s-1 12个梯度,每个梯度下控制测定时间为200 s。仪器自动记录净光合速率(net photosynthetic rate,Pn;μmol·m-2s-1)、光合有效辐射(photosynthetically available radiation,PPFD;μmol·m-2s-1)、蒸腾速率(transpiration rate,Tr;mmol·m-2s-1)等生理参数。叶片水分利用效率(water use efficiency,WUE;μmol·mmol-1)用公式WUE=Pn/Tr计算(Centritto et al., 2000)。
将维持苗木光合速率、蒸腾速率及水分利用效率在最高水平的水分条件作为其最佳的水分环境。绘制光合作用的光响应曲线(Pn-PPFD曲线),依据非直角双曲线模型(Graham,2001)拟合Pn-PPFD的曲线方程,求出最大净光合速率Pn,即光合能力。根据实测数据点的走势估计光饱和点(LSP,μmol·m-2s-1)(Wallin et al., 1992);并对Pn-PPFD曲线的初始部分(PPFD < 200 μmol·m-2s-1)进行线性回归,求得光补偿点(LCP,μmol·m-2s-1)、暗呼吸速率(Rd,μmol·m-2s-1)和表观量子效率(AQY)。
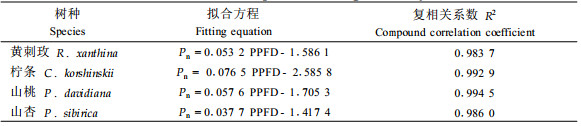
2 结果与分析 2.1 光合能力图 1为4种灌木植物光合速率光响应的模拟值与实测值,可知模拟值与实测值的变化趋势基本一致,模拟方程的复相关系数R2均在0.98以上,说明模型可以较准确地模拟叶片光合速率的光响应规律。本研究结果表明:4种灌木的光合速率对光强的响应规律基本相似,即低光照强度下(PPFD < 300 μmol·m-2s-1),Pn随着光照强度的增加呈正比迅速上升,表明此光强范围内,光强是光合作用的主要限制因子;当超过一定光强,光合速率的增加缓慢。这时叶片来不及吸收和利用高强度的光,同时CO2同化过程的一系列酶促反应速率跟不上而成为光合速率的限制步骤(许大全,2001)。柠条的光合速率在高于LSP后,呈现显著下降的趋势,而其他3种灌木光合速率下降不明显,在LSP之前,相同光照强度下,柠条的光合速率水平最高。由表 3可知,4种灌木的最大光合能力相差较大,柠条、山桃的光合能力较高,分别为26.4、24.5 μmol·m-2s-1,黄刺玫的光合能力最低为15.5 μmol·m-2s-1。柠条的光合能力相当于黄刺玫的1.7倍,山杏的1.44倍,这与许炳成等(2004)研究一致,柠条具有高光合能力的生理特性。

|
图 1 4个树种净光合速率光响应的实测值与拟合曲线 Figure 1 Observed and simulated light response curves of net photosynthetic rates (Pn) of four tree species |
|
|

图 2为PPFD在200 μmol·m-2s-1以下光合速率的拟合曲线,通过回归方程求得表观量子效率、暗呼吸速率及光补偿点(表 4、表 5)。4个树种的复相关系数均在0.98以上,说明方程能够较准确地模拟弱光下光合速率的光响应规律。黄刺玫和山桃的表观光量子效率比较接近,分别为0.053 2和0.056 7,说明它们在利用弱光的能力方面相似。柠条的表观光量子效率最高,为0.076 5,由此可见,柠条在光合能力和对弱光的利用能力方面明显高于其他树种,柠条光合同化潜力最大,对弱光的利用能力强,弱光条件下仍能维持较高的光合能力。山杏的表观量子效率最低,为0.037 7。

|
图 2 弱光下4个树种叶净片光合速率的拟合曲线 Figure 2 Fitting curves of net photosynthstic rates of four tree species in low light intensities |
|
|
|
|
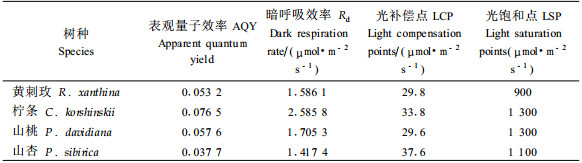
4种灌木的暗呼吸速率表现出一定的差异,黄刺玫、山桃及山杏的暗呼吸速率较为接近,在1.42~1.70 μmol·m-2s-1之间,柠条的暗呼吸速率最高在2.59 μmol·m-2s-1左右。表明在适宜的水分条件下,不同树种对光合产物的消耗能力各不相同。柠条对光合产物消耗的较大,而黄刺玫、山桃及山杏较低的暗呼吸速率可减少呼吸作用对光合产物的消耗,同样有利于干物质的积累。
植物叶片光合作用的饱和光强与光补偿点反映植物对光照条件的要求,是判断植物有无耐荫性的重要指标。研究结果表明:山杏的光补偿点最高为37.6 μmol·m-2s-1,其次为柠条,黄刺玫和山桃较为接近,分别为29.8,29.6 μmol·m-2s-1。说明在适宜的水分条件下,黄刺玫和山桃对弱光的利用方面要高于柠条和山杏,耐荫性较高。山杏对弱光的利用能力最差,这与前面的结果相一致。从光饱和点来看,4种灌木的光饱和点均在900 μmol·m-2s-1以上,柠条和山桃的光饱和点能够达到1 300 μmol·m-2s-1,4种灌木适应较高的光照强度,向阳喜光。
2.3 蒸腾速率植物蒸腾量随土壤水分供给增加而增加(廖建雄等,2002;阮晓等,2005)。当土壤供水充分时,影响蒸腾强弱的环境因子主要是气象因子(郭连生等,1992),蒸腾速率与气象因子的相关性大小依次为光照强度、气温、相对湿度和大气水势(王孟本等,1999)。在适宜水分下,4个树种蒸腾速率的大小顺序为:柠条>黄刺玫>山桃>山杏,在同一光照条件下(PPFD为1 400 μmol·m-2s-1),蒸腾速率由(6.5±0.24)mmol·m-2s-1(山杏)增加到(13.7±0.32)mmol·m-2s-1(柠条),增加幅度在2倍以上,差异很显著(图 3)。4个树种对光照强度的响应规律有所差异,柠条的蒸腾速率随着光照强度的增加而增加,这与郭连生等(1992)的研究一致。在测定的光照强度范围内,柠条的蒸腾速率可达到16.2 mmol·m-2s-1。黄刺玫、山桃和山杏的蒸腾速率随光照强度的增加变化不明显,也就是说土壤水分充足可以维持植物保持一定的蒸腾速率,气孔的开放程度不再增大,蒸腾速率与气象因子的相关性不明显,这与王孟本等(1999)的研究有所差异,这可能与植物自身的特性有关,其原因还需做进一步的研究。

|
图 3 适宜水分下4个树种蒸腾速率的光响应 Figure 3 Light responses of transpiration rates(Tr) of four tree species in optimal water condition |
水分利用效率(WUE)是指植物消耗单位水量生产出的同化量,通常用叶片的净光合速率与蒸腾速率的比值来表示(许大全,2001)。本研究4个树种出现水分利用效率最大值的土壤水分条件分别为50.4%、46.5%、48.6%、51.5%。由图 4可以看出,4个树种的水分利用效率的光响应规律在低光照强度下(PAR < 400 μmol·m-2s-1)基本相似,超过一定的光照强度(光饱和点),柠条出现缓慢下降的趋势,其他树种的变化不明显,也就是说在适宜的水分条件下,光照强度对黄刺玫、山桃及山杏的影响不大,能够适应较宽的光照范围。在同一光照条件下,4个树种的WUE存在明显的差异,山桃的WUE最高,其次为山杏,黄刺玫最低。黄刺玫的WUE与山桃相比,下降幅度在65%左右,这可能与黄刺玫的高蒸腾、低光合特性有关。

|
图 4 4个树种水分利用效率对光照强度的响应 Figure 4 Responses of water use efficiency (WUE) to light intensity of four tree species |
植物的光合能力是用来表征不同植物或者作物品种的重要生理指标(Larcher, 1980)。在最佳或者最适的环境条件下,叶片的最大光合速率表示叶片的最大光合能力(Tartachnyk et al., 2004)。研究表明:在非逆境、饱和光和适宜温度下,不同种植物之间叶片的最大光合速率有很大差异。本研究4种灌木的光合能力介于乔木(贺康宁等,2003;Iryna et al., 2004;Kitaoka et al., 2004)和C4(Jones,1983;Chandrasekar et al., 2000)植物之间,与C3草本(Riepma,1965;Yiruhan et al., 2005)的光合能力相差不大。造成植物光合能力差异的原因可能是其自身体制的自我调节能力不同造成的,也可能是受所处环境的影响(Meir et al., 2002)。植物叶片可以通过多种途径调节光合能力(Le Roux et al., 1999;Warren et al., 2001;Walcroft et al., 2002),以适应外界光环境的变化,这种调节能力是导致种间光合能力差异的主要原因。同时,作为经济型灌木,能更充分地利用捕光色素所吸收的光能来获得较高的光合能力,从而提高其光能利用效率,为碳同化积累更多的能量(ATP)和还原能力(NADPH),为积累更多的光合产物和高产奠定基础。
在一定条件下,表观量子效率反映叶片对光能的利用情况,尤其是对弱光的利用能力(Llusià et al., 2005)。本研究应用非直角双曲线模型较好的模拟4个树种的光响应曲线,所得到的山杏表观量子效率(AQY),与一般植物AQY实验值(0.03~0.05)(许大全,2001)下限接近,黄刺玫及山桃的AQY与许大全(2001)报道的C3植物取值范围(约0.055)基本接近,柠条的表观量子效率最高为0.076 5,说明在适宜的水分条件下,不同植物对弱光的利用能力不同,柠条对弱光的利用能力要高于其他3种被测试植物。同时在水分胁迫或者渍水条件下都会导致4个树种AQY的降低,这与Centritto等(2000)研究是一致的,王孟本等(1999)认为连续降雨植物的AQY也降低,可见适宜的水分条件在很大程度上能够提高植物的表观量子效率。
植物呼吸作用是植物氧化碳水化合物、脂肪、蛋白质等底物生成ATP,CO2和水分的过程,是与光合作用相逆反的过程。虽然暗呼吸消耗植物光合作用中的大部分碳和所固定的能量,但由于这是植物生长所必需的,所以暗呼吸是作为植物正常生长发育的物质和能量源泉,在植物生理生态研究中占据重要位置(Tjoelker et al., 1999;2001)。在适宜的水分和光照条件下,在获得最大净光合速率时,不同树种对光合产物的消耗表现出较大的差异。本研究表明:4个灌木树种的暗呼吸速率在1.42~2.59 μmol·m-2s-1左右,略低于经济型乔木(接玉玲等,2001;Le Roux et al., 1999),明显高于一般的乔木树种(Warren et al., 2001),说明不同树种对环境影响的适应性差异较大,对弱光环境做出的生理学适应各不相同。一般认为植物光补偿点低于20 μmol·m-2s-1,光饱和点低于500 μmol·m-2s-1,为典型阴性植物。而植物的光补偿点在50 μmol·m-2s-1以上,光饱和点在800 μmol·m-2s-1以上,则为典型的阳性植物(Larcher,1980)。与其他研究比较(Riepma,1965;Yiruhan,et al,2005;温达志等,2000),4种植物的光补偿点与光饱和点不同表明山杏具有一定的喜光特性,黄刺玫、柠条和山桃具有较强的适应弱光的能力,对光照环境具有较宽的适应范围。
蒸腾作用在植物水分代谢中起着很重要的调节作用,它的强弱主要取决于土壤中可利用的水分、所必需的能量以及叶片内外间存在的水汽压(水势)梯度,即受外界因子的影响,也受植物体内部结构和生理状况的调节(王孟本等,1999;Iryna et al., 2004)。蒸腾速率反映的是树木单位叶面积瞬间的耗水量,同时可以反映树种调节自身水分损耗能力及适应干旱环境的不同能力,在土壤水分不受限制的条件下,由蒸腾速率的差异可知不同树种的耗水潜力不同(郭连生等,1992;Noble,1999)。本研究结果表明:柠条的耗水潜力最大,瞬时蒸腾耗水速率平均为(12.4±1.78) mmol·m-2s-1,这与王孟本等(1999),许炳成等(2004)研究是一致的,柠条具有深扎根系和较强利用土壤水分的能力,无论在旱季或湿季均保持高光合、高蒸腾速率的光合生理特征。在土壤水分充足时,黄刺玫、山桃及山杏的耗水潜力是由自身的物种特性决定的,受外界环境的影响不明显。它们的耗水能力要明显小于乔木树种及草本植物(郭连生等,1992;接玉玲等,2001;郭志华等,2003),而与一些沙生灌木树种(廖建雄等,2002;阮晓等,2005)比较接近,说明4种灌木树种具有较强的避旱能力,以较低的耗水特性来抵御干旱胁迫。
植物水分利用效率(WUE)是评价植物对环境适应的综合生理生态指标,也是确定植物体生长发育所需水分供应的重要指标之一(许大全,2001)。节水灌溉目的在于最有效地利用水资源,作物的WUE是一个有效的灌溉指标(Turner,1990)、CO2浓度、降低大气湿度(接玉玲等,2001)、土壤控水等措施,均能提高叶片水分利用效率。Turner(1990)曾提出适度水分亏缺有利于作物增产的观点,事实上轻度水分亏缺也能提高植物叶片WUE。本试验结果表明:4种灌木的水分利用效率均在中度水分胁迫下(RWC为50%)达到最大值,这与接玉玲等(2001)的研究是基本一致的。轻度水分亏缺会限制叶片生长,降低Tr,但叶绿体光合酶活性、羧化效率无明显变化,Pn下降很小,水分利用效率较高。
综合上述结果:山桃,山杏虽具有较低的光合能力,但从黄土丘陵沟壑区的水分环境条件来考虑山桃、山杏在该地区具有较高的发展潜力,而且生产上早已发现适度水分亏缺对促进花芽分化、提高果实品质均有良好效果。柠条具有高光合能力、高蒸腾、高水分利用效率的生理特性,并且具有较强的御旱性和耐旱性,因此柠条仍是黄土高原半干旱区最适宜于人工造林的优良灌木树种之一,黄刺玫光合生产能力及有效利用水分方面明显低于柠条、山桃及山杏,在该地区不易进行大面积的人工栽植。
安韶山, 黄懿梅. 2006. 黄土丘陵区柠条林改良土壤作用的研究. 林业科学, 42(1): 70-74. |
程积民, 万惠娥, 杜峰. 2001. 黄土高原半干旱区退化灌草植被的恢复与重建. 林业科学, 37(4): 50-57. |
程积民, 万惠娥, 王静. 2003. 黄土丘陵区山桃灌木林地土壤水分过耗与调控恢复. 土壤学报, 40(5): 691-696. DOI:10.3321/j.issn:0564-3929.2003.05.008 |
郭连生, 刘亮. 1992. 9种阔叶幼树的蒸腾速率, 叶水势和环境因子关系的研究. 生态学报, 12(1): 47-52. DOI:10.3321/j.issn:1000-0933.1992.01.014 |
郭志华, 臧润国, 奇文清, 等. 2003. 两种原始兰科植物生理生态特征的比较. 林业科学, 39(1): 50-55. |
贺康宁, 张光灿, 田阳, 等. 2003. 黄土半干旱区集水造林条件下林木生长适宜的土壤水分环境. 林业科学, 39(1): 10-16. DOI:10.3321/j.issn:1001-7488.2003.01.002 |
胡良军, 绍明安. 2002. 黄土高原植被恢复的水分生态环境研究. 应用生态学报, 13(8): 1045-1048. DOI:10.3321/j.issn:1001-9332.2002.08.027 |
接玉玲, 杨洪强, 崔明刚, 等. 2001. 土壤含水量与苹果叶片水分利用效率的关系. 应用生态学报, 12(3): 387-390. DOI:10.3321/j.issn:1001-9332.2001.03.016 |
李世东, 吴转颖. 2002. 中西部地区退耕还林还草模式探讨. 林业科学, 38(3): 154-159. DOI:10.3321/j.issn:1001-7488.2002.03.027 |
廖建雄, 王根轩. 2002. 干旱、CO2和温度升高对春小麦光合、蒸发蒸腾及水分利用效率的影响. 应用生态学报, 13(5): 547-550. DOI:10.3321/j.issn:1001-9332.2002.05.008 |
牛西午, 丁玉川, 张强, 等. 2003. 柠条根系发育特征及有关生理特性研究. 西北植物学报, 23(5): 860-865. DOI:10.3321/j.issn:1000-4025.2003.05.029 |
阮晓, 王强, 许宁一, 等. 2005. 白梭梭同化枝对干旱胁迫的生理生态响应. 林业科学, 41(5): 28-32. DOI:10.3321/j.issn:1001-7488.2005.05.005 |
孙志蓉, 王文全, 翟明普. 2006. 种间竞争对甘草柠条生物量积累和分配的影响. 林业科学, 42(6): 1-6. |
唐海萍, 史培军. 2001. 沙坡头地区不同配置格局油蒿和柠条水分生态位适宜度研究. 植物生态学报, 25(1): 6-10. DOI:10.3321/j.issn:1005-264X.2001.01.002 |
万雪琴, 胡庭兴, 张健, 等. 2005. 坡耕地退耕还林后的植被恢复. 林业科学, 41(2): 191-194. |
王克勤, 王斌瑞. 1998. 集水造林防止人工林植被土壤干化的初步研究. 林业科学, 34(4): 14-21. DOI:10.3321/j.issn:1001-7488.1998.04.003 |
王孟本, 李洪建, 柴宝峰, 等. 1999. 树种蒸腾作用, 光合作用和蒸腾效率的比较研究. 植物生态学报, 23(5): 401-410. DOI:10.3321/j.issn:1005-264X.1999.05.003 |
王淼, 代力民, 姬兰柱, 等. 2001. 长白山阔叶红松林主要树种对干旱胁迫的生态反应及生物量分配的初步研究. 应用生态学报, 12(4): 496-500. DOI:10.3321/j.issn:1001-9332.2001.04.004 |
汪天, 许成林, 何云核. 2000. 黄刺玫插穗生根能力及其机理的探讨. 安徽农业大学学报, 27(3): 221-224. DOI:10.3969/j.issn.1672-352X.2000.03.004 |
温达志, 叶万辉, 冯惠玲, 等. 2000. 外来入侵杂草薇甘菊及其伴生种基本光合特性的比较. 热带亚热带植物学报, 8(2): 139-146. DOI:10.3969/j.issn.1005-3395.2000.02.010 |
许炳成, 山仑. 2004. 半干旱黄土丘陵区沙棘和柠条水分利用与适应性特征比较. 应用生态学报, 15(11): 2025-2028. DOI:10.3321/j.issn:1001-9332.2004.11.007 |
许大全. 2001. 光合作用效率. 北京: 科学出版社, 13-15.
|
邹厚远, 张信. 1995. 关于黄土高原植被恢复的生态学依据探讨. 水土保持学报, 9(4): 1-4. DOI:10.3321/j.issn:1009-2242.1995.04.003 |
Centritto M, Loreto F, Massacci A, et al. 2000. Improved growth and water use efficiency of cherry saplings under reduced light intensity. Ecological Research, 15(4): 385-392. DOI:10.1046/j.1440-1703.2000.00359.x |
Chandrasekar V, Sairam R K, Srivastava G C. 2000. Physiological and biochemical responses of Hexaploid and Tetraploid wheat to drought stress. J Agronomy & Crop Science, 185(4): 219-227. |
Graham D F. 2001. Models of photosynthesis. Plant Physiology, 125: 42-45. DOI:10.1104/pp.125.1.42 |
Jones H J. Plant and microclimate: A quantitative approach to environmental plant physiology. Cambridge: Cambridge University Press, 134-135.
|
Kiefer J C, Wolfowitz J. 1959. Optium design in regression problem. Ann Math Statist, 30: 271-294. DOI:10.1214/aoms/1177706252 |
Kitaoka S, Koike T. 2004. Invasion of broad-leaf tree species into a larch plantation: seasonal light environment, photosynthesis and nitrogen allocation. Physiologia Plantarum, 121(4): 604-611. DOI:10.1111/ppl.2004.121.issue-4 |
Larcher W. 1980. Physiological Plant Ecology. Beijing: Academic Press.
|
Le Roux X, Grand S, Dreyer F, et al. 1999. Parameterization and testing of a biochemically based photosynthesis model for walnut (Juglans regia) trees and seedlings. Tree Physiol, 19: 481-492. DOI:10.1093/treephys/19.8.481 |
Llusià J, Peñuelas J, Munné-Bosch S. 2005. Sustained accumulation of methyl salicylate alters antioxidant protection and reduces tolerance of holm oak to heat stress. Physiologia Plantarum, 124(3): 353-361. DOI:10.1111/ppl.2005.124.issue-3 |
Meir P, Kruijt B, Broadmeadow M, et al. 2002. Acclimation of photosynthetic capacity to irradiance in tree canopies in relation to leaf nitrogen concentration and leaf mass per unit area. Plant, Cell & Environment, 25(3): 343-357. |
Noble P S. 1999. Physiochemical and environment plant physiology. (Second Edition). San Diego: Academic Press, 293-349.
|
Riepma P. 1965. The effect of disodium methylarsonate, dalapon, amitrole-T and paraquat on Axonopus Compressus, ottochloa no dosa and Paspalum Conjugatum. Weed Research, 5(2): 151-157. DOI:10.1111/wre.1965.5.issue-2 |
Tartachnyk I I, Blanke M M. 2004. Effect of delayed fruit harvest on photosynthesis, transpiration and nutrient remobilization of apple leaves. New Phytologist, 164(3): 441-450. DOI:10.1111/j.1469-8137.2004.01197.x |
Tjoelker M G, Oleksyn J, Lee T D, et al. 2001. Direct inhibition of leaf dark respiration by elevated CO2 is minor in 12 grassland species. New Phytologist, 150(2): 419-424. DOI:10.1046/j.1469-8137.2001.00117.x |
Tjoelker M G, Reich P B, Oleksyn J. 1999. Changes in leaf nitrogen and carbohydrates underlie temperature and CO2 acclimation of dark respiration in five boreal tree species. Plant, Cell & Environment, 22(7): 767-778. |
Turner N C. 1990. Plant water relation and irrigation management. Agric Water Manager, 17: 59-73. DOI:10.1016/0378-3774(90)90056-5 |
Walcroft A, Le Roux X, Diaz-espejo A, et al. 2002. Effects of crown development on leaf irradiance, 1eaf morphology and photosynthetic capacity in a peach tree. Tree Physiol, 22: 929-938. DOI:10.1093/treephys/22.13.929 |
Wallin G, Skärby L, Selldén G. 1992. Long-term exposure of Norway spruce, Picea abies (L.) Karst., to ozone in opentop chambers. Ⅲ. Effects on the light response of net photosynthesis in shoots of different ages. New Phytologist, 121(3): 387-394. DOI:10.1111/nph.1992.121.issue-3 |
Warren C R, Adama M A. 2001. Distribution of N, Rubisco and photosynthesis in Pinus pinaster and acclimation to light. Plant Cell Environ, 26(4): 597-609. |
Yiruhan S, Shiyomi M, Takahashi S, et al. 2005. Evaluating the adaptability of herbage species to environmental variation through a long-term grazing experiment. Grassland Science, 51(4): 287-295. DOI:10.1111/grs.2005.51.issue-4 |