 2008, Vol. 44
2008, Vol. 44文章信息
- 康东东, 韩利慧, 马鹏飞, 魏学智, 毕润成.
- Kang Dongdong, Han Lihui, Ma Pengfei, Wei Xuezhi, Bi Runcheng
- 不同地理环境下酸枣叶的形态解剖特征
- Comparison on Characters of Leaf Anatomy of Ziziphus jujuba var. spinosa in Different Geography Environment
- 林业科学, 2008, 44(12): 135-139.
- Scientia Silvae Sinicae, 2008, 44(12): 135-139.
-
文章历史
- 收稿日期:2007-11-09
-
作者相关文章



水资源短缺是目前制约农业生产的主要问题,全球约有43%的耕地为干旱、半干旱地区。在自然条件下,干旱是决定植被地理分布和限制作物产量的主要因素(Schulze,1986)。提高植物的抗旱能力已成为现代植物研究中亟需解决的关键问题之一。干旱、半干旱地区的植物长期处于干旱的影响下,在形态、解剖构造和生理功能上均发生一系列适应性变化,构成植物的抗旱性(Kramer et al., 1979)。叶是光合作用、气体交换及蒸腾最重要的位点,与周围环境有密切联系,其形态结构对生境的变化反应最为敏锐,可塑性也最大。因而,植物对不同环境的适应性改变易反映在叶的形态结构上(Wylie,1951;Jackson,1967)。
酸枣(Ziziphus jujuba var. spinosa)为鼠李科枣属的一种灌木或小乔木,是具有较高经济价值的野生果树资源,也是重要的药用植物,还是防风固沙、保持水土的先锋树种。酸枣主要分布于我国北方地区,其生态幅很广,从湿润的东海海滨到极度干旱的新疆均能正常生长。目前对植物抗旱性的研究主要集中在小范围生境内(崔秀萍等,2006;安黎哲等,2004),而对同一种植物在跨经度大范围生境中结构变异规律的报道却很少(StHilsire et al., 1999)。小范围生境变化对植物生长的影响有限,而小范围生境差异与大范围生境差异对植物生长造成的影响,对解决干旱的机制决定植被地理分布问题有重要意义。本文对不同经度的酸枣叶结构解剖观察,从叶的解剖结构上探索其抗旱机制,阐释植物形态结构和大范围环境变化的相互适应机制及规律,为黄土高原地区开发利用酸枣资源提供科学依据。
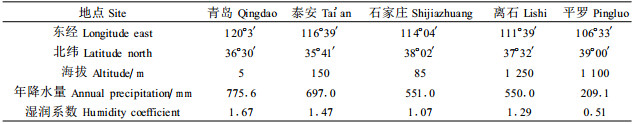
1 材料与方法 1.1 试验材料酸枣叶分别采自山东青岛、山东泰安、河北石家庄、山西离石、宁夏平罗。采样地点概况见表 1。
|
|
试验材料均取自酸枣分布地向阳山坡处,随机取胸径为5 cm左右的植株5株,选取其向阳面大枝的中部成熟叶进行采集,采样后立即用FAA固定。试验时每采集地分别从所采5株植株叶片中每植株随机选取1个叶片进行制样,每个制样取10个视野的平均值,每采样地共得5个平均值。主要制样方法:1)常规石蜡切片:切片厚12 μm,番红固绿对染,NiKon E200光学显微镜下观测照相;2)叶表皮处理方法:取中部叶脉附近的部分切成约1 cm×1 cm小块,FAA固定24 h以上,转移到20%次氯酸钠溶液中,在35 ℃恒温箱中离析6~8 h,撕取表皮,在NiKon E200光学显微镜下观测照相;3)扫描电镜制样:取中部叶脉处,剪成约5 mm×5 mm小块,FAA固定24 h以上,70%的酒精洗涤,并逐级脱水,醋酸异戊酯过渡,CO2临界点干燥,干燥后的样品粘在样品台上并镀金处理,在KYKY2800扫描电镜观察拍照。所测数据用SPSS13.0软件进行方差分析和LSD(最小显著差数法)多重比较。
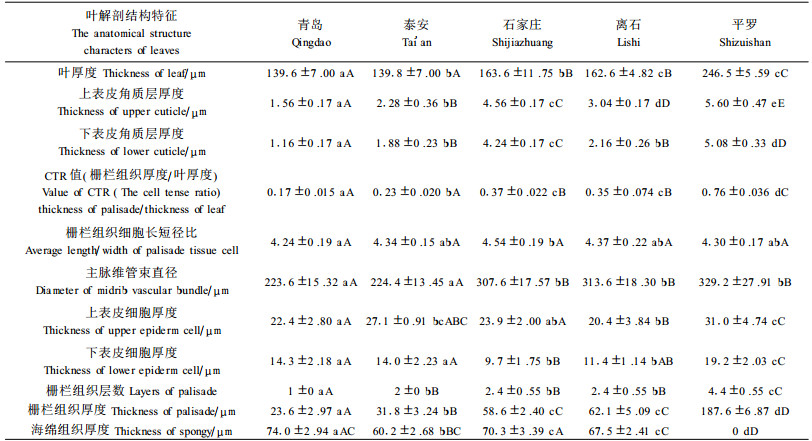
2 结果与分析 2.1 叶横切面结构1) 不同生境酸枣叶解剖结构的共同特征 叶的横切面(图版Ⅰ-16~20),酸枣上、下表皮各由一层细胞组成,细胞排列紧密,呈矩圆形或矩形,上、下表皮细胞外壁均被角质层,上表皮细胞及其角质层均较下表皮厚,叶肉均包括栅栏与海绵组织。主脉向下突起、半圆形,是一个外韧维管束(图版Ⅰ-11~15)。

|
图版Ⅰ Plate Ⅰ 1-20:光镜下 Under light microscope;21-35:电镜下 Under scanning electron microscope. 1-5:青岛、泰安、石家庄、离石、平罗(下同),上表皮细胞(单位标尺30 μm) Upper epidermis cells (the unit scale30 μm) in Qingdao,Taian,Shijiazhuang,Lishi,Pingluo (The same below);6-10:下表皮细胞(单位标尺30 μm) Lower epidermis cells(the unit scale 30 μm);11-15:主脉(单位标尺40 μm) Aremidrib (the unit scale 40 μm);16-20:横切面(单位标尺30 μm) Transverse section (the unit scale 30 μm);21-25:上表皮表面角质层纹式(单位标尺20 μm) Upper cuticle ornamentation (the unit scale30 μm);26-30:下表皮表面角质层纹式(单位标尺30 μm) Lower cuticle ornamentation (the unit scale30 μm);31-35:下表皮表面气孔(单位标尺5 μm) Stomata in proximal surface (the unit scale5 μm). |
2) 不同生境酸枣叶解剖结构的差别 叶的横切面(图版Ⅰ-16~20),青岛到平罗随着经度的变化,酸枣叶厚度、上表皮角质层厚度、下表皮角质层厚度、上表皮细胞厚度、下表皮细胞厚度呈增加的趋势;但分布于离石的酸枣叶厚度、上表皮角质层厚度、下表皮角质层厚度、上表皮细胞厚度几个指标较分布于石家庄的酸枣低。随着经度的变化酸枣叶的栅栏组织层数、栅栏组织厚度、主脉维管束直径、CTR值均升高,海绵组织层数则相反,栅栏组织细胞长短径比(栅栏组织细胞系数)变化不大。海绵组织细胞长短径比则随着经度的降低,细胞形状逐渐由长圆形变为长柱形,细胞间隙逐渐变小,细胞排列变紧密,转化为栅栏组织。叶逐渐由异面叶转变为等面叶(表 3)。
|
|
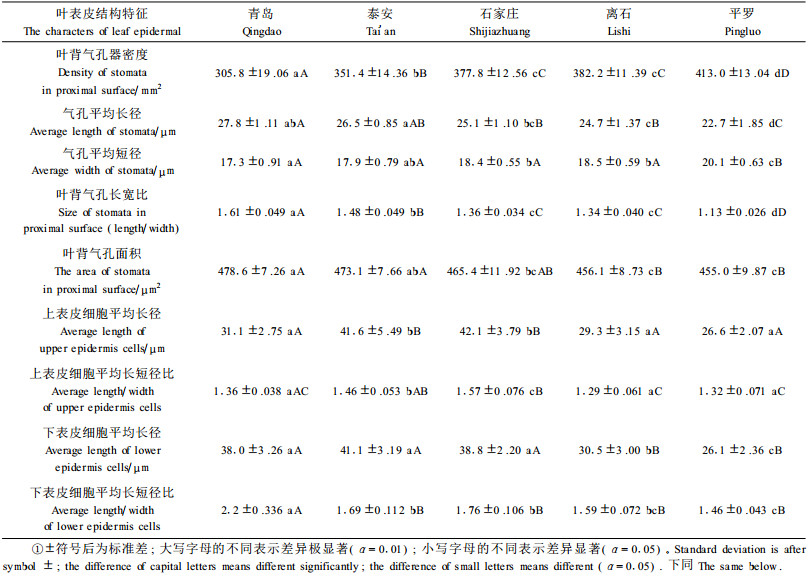
1) 光镜下的结构特征 叶的表面观,不同产地酸枣叶的气孔类型均为不规则形,形状稳定,不随生境条件变化而变化。气孔均分布在下表皮(图版Ⅰ-6~10)。从青岛到平罗随着分布地经度的变化,气孔密度从305.8个·mm-2个增加至413.0个·mm-2,气孔面积与气孔长宽比逐渐变小;而上表皮细胞形状与大小变化未表现出规律性,其形状有矩形、近似矩形、五边形、六边形分布不等,上表皮细胞以石家庄为最大,其长径达42.1 μm,长短径比达1.57,以平罗为最小,其长径为26.6 μm,长短径比为1.32(表 2)。随着经度的变化,下表皮细胞长短径比逐渐变小,形状逐渐由长条形细胞居多转变为以五、六边形居多。下表皮细胞以泰安为最大,其长径达41.1 μm,长短径比达1.69,以平罗为最小,其长径为26.1 μm,长短径比为1.46(表 3)。上下表皮细胞垂周壁样式均为平直型。青岛、泰安、石家庄、离石4地样品的表皮毛均稀疏地分布在上下表皮的主脉上,而平罗采集的酸枣叶的表皮毛密集地分布在主脉。表皮毛均为多细胞型。
|
|
2) 扫描电镜下叶的结构特征 从青岛到平罗随着经度和湿润系数的变化,酸枣叶上表皮纹饰变化极显著,表皮细胞轮廓由清晰逐渐变得不可分辨。青岛样品表皮表面平整,看不到角质纹饰,沿胞间隙处呈规则的线状凹陷,可清晰地看到五边形和六边形的表皮细胞(图版Ⅰ-21)。泰安样品上表皮表面稍有凹凸起伏,沿表皮细胞垂周壁处形成条状凸起,但在胞间隙处仍呈凹陷,仍可分辨出细胞的形态(图版Ⅰ-22)。石家庄样品上表皮形成极明显的不规则网状纹饰,已看不清细胞的轮廓(图版Ⅰ-23)。离石样品上表皮纹饰形状与石家庄纹饰相似,但形状较规则(图版Ⅰ-24)。平罗样品上表皮为明显凸起的山包状纹饰,且向外辐射出条状纹饰相互连接成不规则的网状(图版Ⅰ-25)。以上5个样地叶表面纹饰的变化有明显规律,即:平滑-条状—网状—山包状。网线为表皮细胞垂周壁处的凸起,网眼部分为表皮细胞平周壁处,山包状的结点为3个细胞或4个细胞的接合处。这是因为表皮细胞垂周壁分泌的角质层在表皮表面的细胞接合处集中堆积,形成不同形状的纹饰。
从青岛到平罗随着经度的变化,酸枣叶下表皮表面纹饰变化无明显的变化规律。青岛的样品下表皮表面较平整,有稀疏的条状纹饰,呈不规则的分布。泰安的样品,纹饰明显但杂乱无规则。石家庄样品表皮纹饰杂乱,略显网状,网线较粗,网眼小形状不规则。离石的样品则为较规则网状纹饰。平罗样品下表皮表面纹饰分布无有明显规律,可见山包状凸起,无明显的网格。随着经度的变化,气孔与表皮细胞的相对位置发生变化,平罗酸枣叶气孔凹陷,其余基本与表皮相平(图版Ⅰ-26~35)。气孔长径逐渐变小,由青岛27.8 μm增加为平罗22.7 μm(表 2)。
2.3 抗旱指标的数据统计分析植物对干旱环境的适应经历着不同的方式和途径,这些特征在处于不同生境的植物中表现的差异程度不一致。因此对一系列指标进行了描述性分析与多重比较分析(见表 2、3)。
方差分析结果可知(表 2、表 3):生长在不同生境的酸枣除了栅栏组织长短径比指标外其他指标均出现极显著差异。全部指标灵敏度大小顺序依次是:上表皮角质层厚度>叶背气孔器密度、叶背气孔长宽比、下表皮角质层厚度、栅栏组织厚度、海绵组织厚度>气孔平均长径、上表皮细胞平均长短径比、叶厚度、CTR值、上表皮细胞厚度、下表皮细胞厚度、栅栏组织层数>主脉维管束直径、气孔平均短径、叶背气孔面积、下表皮细胞平均长径、下表皮细胞平均长短径比>栅栏组织细胞长短径比。
3 结论与讨论植物器官的形态结构与其生理功能和生长环境密切相适应。长期在外界生态因素的影响下,叶形态结构的变异性和可塑性最大,即叶对生态条件的反映最为明显(朱弧,1993)。我国北方地区影响植物生长的主要限制因子是水分,由于青岛、泰安、石家庄、离石和平罗5地湿润系数或降水量的变化,导致酸枣叶结构一系列的梯度变化。青岛、泰安、石家庄、离石、平罗5地均处于(37±2)°N内。青岛位于胶东半岛,濒临黄海;泰安位于山东省中部泰山脚下,两地均为暖温带半湿润大陆性季风气候。石家庄位于河北省中南部,西傍太行山;离石位于吕梁山中段西侧,均为暖温带大陆性季风气候。平罗位于宁夏回族自治区北部,光照充足,蒸发强烈,空气干燥,为温带干旱气候区。从青岛到平罗随着经度的变化,5地年平均降水量显著降低,湿润系数依次递减(离石除外),逐渐由半湿润区过渡到干旱地区。
青岛酸枣叶具有典型的中生结构:叶片厚度小,CTR值小,栅栏组织层数少,厚度小,排列疏松,主脉直径小,角质层薄,气孔密度小,气孔位置与表皮相平,气孔长宽比大。平罗酸枣叶则有典型的旱生结构特征,叶片厚度大,CTR值大,栅栏组织层数多,厚度大,主脉直径大,角质层厚,气孔密度大,气孔下陷,气孔长宽比小。而泰安、石家庄、离石为由中生趋于旱生的过渡类型。在大范围生境的变化条件下,一些形态结构特征变化的抗旱机理也得到验证。叶肉细胞变小,叶厚度变大,长短径比增大,叶表面积与体积比变小,有利于叶绿体垂直叶表排列,提高光合作用效率及植物对水分的利用率。栅栏组织层数较多可以提高栅海比,叶肉组织排列紧密有利于光合作用的有效积累,防止强辐射对叶肉的损伤。发达的机械组织可赋予植物叶子坚韧性,从而保证叶肉组织不因叶的失水萎缩变形而受伤害,而发达的输导组织则能保证和提高水分输导效率(李正理等,1981;周智彬等,2002;赵翠仙等,1981)。叶表面覆盖的角质层是一种类脂膜,其主要功能是减少水分向大气散失,是植物水分蒸发的屏障,除有不透水性外,还能抵抗微生物的侵袭(王绍辉等,2003)。厚的角质层可提高植物的能量反射与降低蒸腾,从而增强植物的抗旱性(祁如虎等,1998)。对于气孔密度与抗旱性的关系,一般认为“旱生植物的气孔密度增加是一种适应特征”、“气孔密度增加是植物对旱生环境的适应,气孔密度低是植物对湿生环境的适应”,在环境干旱程度增加时提高气孔分化的程度,增加气孔密度,可能是待水分供应充足时,增加气体交换,以提高光合作用效率(Cutter, 1973)。对于气孔位置与抗旱性的关系,凹陷的气孔可能有利于滞留CO2和H2O,在旱生条件下可为光合作用提供充足的原料,同时还可以使内在空气的饱和差值减小,阻止水分散失,这可能是植物自然产生的抗御蒸腾的机制(胡适宜,1959)。气孔长宽比减小可能是为在长径一定的情况下增加开口度,加大呼吸面积,从而补偿呼吸效率,增加通气量,使酸枣的气孔结构具有较强的抗逆特性(李正理,1981)。从青岛到平罗,叶表面特征由平滑逐渐变为条状、网状、和山包状。上下表皮有相同的变化趋势。凸起的纹饰总是集中分布于表皮细胞的彼此接合处,推测认为凸起的角质层纹饰主要由垂周壁分泌到细胞外面,通过胞间隙在表皮细胞外壁的接合处累积而形成。
对相关指标作的多重比较表明随着经度的变化,由东向西,年平均降水量的逐渐减少,酸枣叶的主要性状均出现适应性结构,从中生植物的结构变为旱生植物的结构。酸枣叶各性状对环境变化的敏感程度是不同的。栅栏组织细胞长短径比指标敏感度最低而CTR值指标敏感度中等说明在适应过程中栅栏组织的增厚主要依靠细胞层数的增加而非细胞形状的变化。表皮细胞的相关指标敏感度为中等或偏低,结合其三维结构的长度观察结果表明:表皮细胞在适应干旱的过程中不仅仅表现为细胞大小的变化而且出现了细胞形状由横长方体到竖长方体的转变,因此单独使用三维细胞的某一维来分析细胞大小的变化会产生一定的片面性,这样的变化使表皮细胞排列更加致密,提高其抗旱性。CTR值、栅栏组织层数、叶厚度、主脉维管束直径的敏感度为中等或偏低说明其会对环境中水分的变化做出响应但较保守,在抗旱育种中参考性较高。角质层表现出最高的敏感度这可能与它在叶结构的最外部有关系。
关于叶表面特征植物分类学意义,对酸枣的研究支持“单纯的植物解剖特征不能用作分类的依据,其被认为是对分类建立时所依据的外部形态学特征的补充与延伸”的论断(Metcalfe et al., 1950);有部分研究将叶表皮微形态学特征作为属内种间分类的依据之一(周守标等,2007;丛义艳等,2007)。对酸枣的研究尤其是对不同地区叶表面纹饰巨大差异的研究使我们对此产生质疑,同时支持陈之端等认为的“叶表皮性状在一定程度上能反映分类群间的系统发生,可用于科下属间关系的探讨”的观点(任辉等,2003)。叶表面特征与生长环境因素之间的相互适应机理尚有待于进一步深入研究。
安黎哲, 戴怡龄, 陈拓, 等. 2004. 乌鲁木齐河源区不同海拔的火绒草叶片结构特征的比较研究. 冰川冻土, 26(4): 474-481. DOI:10.3969/j.issn.1000-0240.2004.04.015 |
丛义艳, 刘克明, 陈薇, 等. 2007. 6种凤仙花属(Impatiens L.)植物叶表皮特征的微形态学研究. 湖南师范大学自然科学学报, 30(1): 68-71. DOI:10.3969/j.issn.1000-2537.2007.01.017 |
崔秀萍, 刘果厚, 张瑞麟. 2006. 浑善达克沙地不同生境下黄柳叶片解剖结构的比较. 生态学报, 26(6): 1842-1847. DOI:10.3321/j.issn:1000-0933.2006.06.027 |
胡适宜. 1959. 植物学(形态学及解剖学部分). 北京: 高等教育出版社.
|
李正理, 李荣敖. 1981. 我国甘肃九种旱生植物同化枝的解剖观察. 植物学报, 23(3): 181-183. |
李正理. 1981. 旱生植物的形态和结构. 生物学通报, (4): 9-13. |
祁如虎, 李有忠, 范建平. 1998. 16种高山植物叶内的异细胞及其生态学意义. 西北植物学报, 18(2): 270-276. DOI:10.3321/j.issn:1000-4025.1998.02.022 |
任辉, 潘开玉, 陈之端, 等. 2003. 葡萄科植物叶表皮特征及其系统学意义. 植物分类学报, 41(6): 531-544. |
王绍辉, 张福墁. 2003. 黄瓜叶表面特性与生态适应性. 生态学报, 23: 199-204. DOI:10.3321/j.issn:1000-0933.2003.01.024 |
赵翠仙, 黄子琛. 1981. 腾格里沙漠主要旱生植物旱性结构的初步研究. 植物学报, 23(4): 278-283. |
周守标, 王影, 李金花, 等. 2007. 安徽贝母属植物的叶表皮形态研究. 中国中药杂志, 32(2): 105-108. DOI:10.3321/j.issn:1001-5302.2007.02.005 |
周智彬, 李培军. 2002. 我国旱生植物的形态解剖学研究. 干旱区研究, 3(1): 35-40. |
朱弧.1993.几种猕猴桃叶解剖结构与抗旱性关系的研究.中国植物学会60周年年会学术论文汇编.北京: 科学出版社, 243-244.
|
Cutter E G.1973.植物解剖学.李正理, 译.北京: 科学出版社.
|
Jackson L W R. 1967. Effect of shade on leaf structure of deciduous tree species. Ecology, 48: 498-499. DOI:10.2307/1932686 |
Kramer P J, Kozlowski T T. 1979. Physiology of Woody Plants. Academic Press Inc.
|
Metcalfe C R, Chalk L. 1950. Anatomy of the Dicotyledons. Oxford: Clarendon Press.
|
Schulze E D. 1986. Carbon dioxide and water vapor response to drought in the atmosphere and in the soil. Annu Rev Plant Physiol, 37: 247-274. DOI:10.1146/annurev.pp.37.060186.001335 |
StHilsire R, Graves W R. 1999. Foliar Traits of Sugar Maples and Black Maples Near 43°N Latitude in the Eastern and Central. United States. J Amer Soc Hort Sci, 124(6): 605-611. |
Wylie R B. 1951. Principles of foliar organization shown by sunshade leaves from ten species of deciduous dicotyledon trees. American Journal of Botany, 38: 355-361. DOI:10.1002/ajb2.1951.38.issue-5 |