 2008, Vol. 44
2008, Vol. 44文章信息
- 李巧, 陈又清, 周兴银, 康海艳, 陈彦林, 吴俊, 郭文俊.
- Li Qiao, Chen Youqing, Zhou Xingyin, Kang Haiyan, Chen Yanlin, Wu Jun, Guo Wenjun
- 云南元谋干热河谷捕食性昆虫多样性
- Diversity of Predatory Insect in Yuanmou Arid-Hot Valley, Yunnan
- 林业科学, 2008, 44(10): 82-87.
- Scientia Silvae Sinicae, 2008, 44(10): 82-87.
-
文章历史
- 收稿日期:2007-01-23
-
作者相关文章


2. 中国林业科学研究院资源昆虫研究所 昆明 650224
2. Research Institute of Resources Insects, CAF Kunming 650224
云南元谋干热河谷位于金沙江一级支流龙川江河谷,土地严重荒漠化,是生态环境严重脆弱的地区。由于长期的不合理开发,植被破坏严重,水土流失加剧,自然灾害发生频繁,云南元谋干热河谷生态环境日趋恶化,生物多样性保护形势严峻。自20世纪80年代以来,元谋干热河谷植被恢复研究日益受到重视,植被恢复过程中昆虫多样性研究也日趋丰富(李巧等,2006a;2006b;2007;李昆等,2006);而元谋干热河谷捕食性昆虫多样性的研究未见报道。
捕食性昆虫在生态系统中具有调节植物和植食生物种群数量的作用,它们保护各种生物共存、防止任何一种生物数量过度丰富,使得地球上物种呈现出多样化(丁建清等,1996)。捕食性昆虫多样性的研究,是生物多样性研究的一个重要组成部分,对于丰富全球生物多样性信息库及发挥生物防治在林业可持续发展中的作用具有重大的理论和实践意义(任炳忠等,2001)。国内的研究更多集中在对农业生态系统捕食性昆虫群落结构与动态的报道,对森林生态系统捕食性昆虫群落的研究十分少见(陈振耀等,2001;林荣华等,2003;侯美珍等,2005;罗淑萍等,2006;师光禄等,2006);而国外的研究已扩展到捕食性昆虫多样性对生态系统功能影响方面(Hart,2002;Deborah et al., 2004)。本文对云南元谋干热河谷捕食性昆虫进行调查,提供捕食性昆虫物种多样性的基本信息,并选取元谋干热河谷分布面积较大的9种生境为研究对象,通过考察各生境捕食性昆虫群落特征,揭示不同生境捕食性昆虫在种类组成和多样性方面的差异,对于认识元谋干热河谷捕食性昆虫资源状况、实现科学保护和合理开发利用具有积极意义。
1 研究方法 1.1 研究地点的选择根据元谋干热河谷植被性质的不同设置9块试验地,进行捕食性昆虫群落多样性研究。人工植被选择造林面积较大的云南松林、桉树林、桉树新银合欢林、桉树印楝林、印楝林及多树种混交的人工林作为试验地;自然植被选择分布较广的牛筋条-黄荆林、余甘子-明油子灌木林及明油子-扭黄茅灌草丛作为试验地。各试验地面积约1 hm2,概况如表 1所示。
|
|

元谋干热河谷捕食性昆虫物种多样性调查在各种类型的生态系统内进行,于2004—2005年运用网捕法、震落法、陷阱法及灯诱法等多种方法采集捕食性昆虫标本,将采集到的标本用75%酒精保存或置于三角纸包内,带回实验室整理,联系有关专家进行种类鉴定。
捕食性昆虫群落调查于2005年1、3、5、7、9及11月进行,应用网捕法采集生活于乔木、灌木及草本层的灌木-草本层捕食性昆虫,应用陷阱法采集地表活动的捕食性昆虫。在每个试验地内设置一定的调查路线, 由2名调查人员使用直径400 mm、深750 mm的拉杆式捕虫网进行定期网捕采集,每次扫网1 h。在每个试验地内设置口径80 mm、高150 mm的1次性塑料杯10个,分为2组:第1组以乙二醇作为诱剂,每个诱杯内倒入50 mL诱剂;第2组以糖醋液作为诱剂,糖醋液为白糖、食醋、酒精及水的混合液, 其比例为1:2:2:20,每个诱杯内倒入80 mL诱剂;每1组诱杯间距10 m, 2组间相距20 m;每个诱杯上方放置好防雨的石板;诱集时间为5 d,标本保存与鉴定同前。
1.3 数据分析群落内多样性(α多样性)通过物种丰富度指数、物种多样性指数和均匀度指数等进行测度(马克平,1994)。物种丰富度指数用S表示, 即物种的数目;物种多样性指数采用Shannon-Wiener指数H′,物种优势度采用Simpson优势度指数C,均匀度指数采用Pielou指数E。群落间的多样性(β多样性)采用Jaccard相似性系数(q值)进行群落相似性测度,根据Jaccard相似性系数原理,当q为0.00~0.25时,为极不相似;当q为0.25~0.50时,为中等不相似;当q为0.50~0.75时,为中等相似;当q为0.75~1.00时,为极相似。本文利用Excel计算H′、C及E,利用EstimateS(Version7.5.0)软件计算q值(Colwell,2005)。
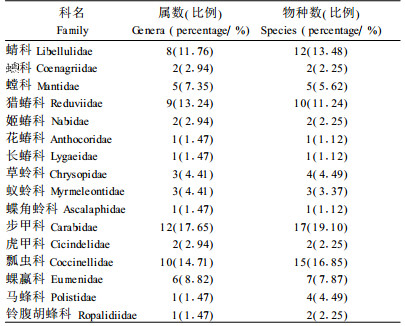
2 结果与分析 2.1 元谋干热河谷捕食性昆虫物种多样性 2.1.1 元谋干热河谷捕食性昆虫名录通过广泛调查,共采集捕食性昆虫89种(含10个待定种),涉及5目、16科、68属,其中中国新记录1种,云南新记录11种(表 2)。
|
|

元谋干热河谷捕食性昆虫属数较多的科有步甲科、瓢虫科、猎蝽科、蜻科、蜾蠃科等,种类丰富的科有步甲科、瓢虫科、蜻科、猎蝽科、蜾蠃科等(表 3),而步甲科、瓢虫科、猎蝽科和蜻科的属及种的比率均超过了10%。可见,这4科是元谋干热河谷捕食性昆虫的优势科,而步甲科占显著优势。
|
|
比较各试验地内捕食性昆虫群落物种丰富度(表 4),大小顺序依次为:Ⅱ>Ⅴ>Ⅰ=Ⅵ>Ⅸ> Ⅲ=Ⅳ=Ⅶ>Ⅷ,即桉树印楝林捕食性昆虫种类最丰富,多树种混交的人工林次之;而桉树林、桉树新银合欢林及牛筋条-黄荆林捕食性昆虫种类较贫乏,余甘子-明油子灌木林最贫乏。从各试验地内捕食性昆虫群落的多样性指数来看,Shannon-Wiener指数值大小顺序依次为: Ⅱ>Ⅴ>Ⅵ>Ⅸ>Ⅲ>Ⅰ>Ⅳ>Ⅶ>Ⅷ;而优势度指数和均匀度指数反映出的结果略有不同。综合这几种指数,桉树印楝林、多树种混交的人工林及印楝林多样性及均匀度指数较高、优势度较低,具有较高的多样性;牛筋条-黄荆林与余甘子-明油子灌木林多样性及均匀度指数较低、优势度较高,具有极低的多样性。桉树林及桉树新银合欢林尽管捕食性昆虫种类均较贫乏,但桉树新银合欢林由于优势度较低、均匀度较高,其多样性高于桉树林。云南松林尽管捕食性昆虫种类较桉树林及桉树新银合欢林丰富,但由于优势度较高、均匀度较低,其多样性位于两者之间。明油子-扭黄茅灌草丛具有比较丰富的捕食性昆虫种类,捕食性昆虫群落优势度较低、均匀度较高,其多样性较高。
|
|

比较各试验地内捕食性昆虫群落之间的相似性,相似性系数q值在0.111~0.500之间,为极不相似至中等相似水平(表 5)。其中,桉树印楝林和印楝林捕食性昆虫群落之间的相似性最大,达到中等相似水平,这与其在乔木、灌木及草本的构成上均有共有种有关;印楝林与明油子-扭黄茅灌草丛、牛筋条-黄荆林之间,桉树印楝林与桉树林、牛筋条-黄荆林之间具有较大的相似性,接近中等相似水平;其余各试验地内捕食性昆虫群落之间的相似性均较低,其中多树种混交的人工林与牛筋条-黄荆林之间相似性最低。
|
|

报道了元谋干热河谷捕食性昆虫89种(含待定种10种),其中中国新记录1种,云南新记录11种。步甲科种类最丰富,占所有捕食性昆虫种类的19.1%;瓢虫科、猎蝽科和蜻科种类较丰富。
捕食性昆虫多样性与其猎物多样性有关,而猎物多样性即植食性昆虫多样性又与寄主植物种类及结构多样性有关;寄主的种类越多,分布范围越广, 天敌的多样性便越大(丁建清等,1996;Longcore,2003)。元谋干热河谷9种生境捕食性昆虫群落多样性按从大到小的排序为:桉树印楝林>多树种混交的人工林>印楝林>明油子-扭黄茅灌草丛>云南松林>桉树新银合欢林>桉树林>牛筋条-黄荆林>余甘子-明油子灌木林。从自然植被的捕食性昆虫群落来看,干热河谷分布最普遍的明油子-扭黄茅灌草丛比牛筋条-黄荆林和余甘子-明油子灌木林具有较高的多样性和均匀性,较低的优势度,依照多样性与稳定性呈正相关理论(Odum,1953;MacArthur,1955;Tilman et al., 1998;Yachi et al., 1999),明油子-扭黄茅灌草丛捕食性昆虫群落较稳定,这种稳定和柴宗新等(2001)认为明油子-扭黄茅灌草丛是顶级群落的观点相吻合;而明油子-扭黄茅灌草丛捕食性昆虫群落较高的多样性和均匀性应得益于其生境面积的绝对优势。牛筋条-黄荆林和余甘子-明油子灌木林草本层和地被物不发达,加之政府或企业的开山修路、采石等行为,生境片段化较严重,它们在干热河谷的分布面积也日益缩小;生境片段化阻碍包括捕食性昆虫在内的生态系统的基因交流,而日益缩小的生境可能会导致这些分布面积不广泛的自然植被的严重退化甚至最终消失。在人工植被中,封育时间近20年的桉树印楝林捕食性昆虫群落具有最高的多样性,反映出近20年的人工封育措施对多样性保护起到了积极作用;封育时间仅5~7年多树种混交的人工林及印楝林捕食性昆虫群落具有较高的多样性;桉树新银合欢林和桉树林具有较低的多样性,云南松林多样性居中。李巧等(2006a;2006b)对元谋干热河谷直翅目和鞘翅目昆虫多样性研究显示:植被恢复初期,由于栖境改善和食料的增加,直翅目和鞘翅目群落处于建立和发展阶段,种类和数量增加,群落多样性提高。因此,多树种混交的人工林及印楝林捕食性昆虫群落具有较高的多样性应该是由于其植食性昆虫群落多样性在植被恢复初期增加而导致捕食性昆虫群落多样性增加的缘故。桉树新银合欢林和桉树林封育时间和桉树印楝林相同,但其捕食性昆虫群落多样性极低,这是由于其草本层极不发达、地表裸露及植被组成简单。
从群落相似性来看,一些人工植被类型与自然植被类型捕食性昆虫群落之间接近中等相似水平,如印楝林与明油子-扭黄茅灌草丛、桉树印楝林与牛筋条-黄荆林等,反映出这些人工植被类型的建立,是在原有的自然植被上种植乔木,而这些乔木树种尚未成为影响植物群落发展的优势种,无法主导植物群落的演替,因而表现在捕食性昆虫群落上亦未形成独特的群落。至于这些乔木树种是否能主导植物群落的演替,是制定干旱河谷生态恢复策略所要求回答的。如果这些乔木树种能够主导植物群落的演替,那么多长时间能实现演替,还需要进一步调查;如果不能,则表明,植树造林的干旱河谷生态恢复策略是不切实际的,而这种观点与Moseley等(2006)的研究相一致。
封山育林和退耕还林还草政策的实施,在很大程度上保护了元谋干热河谷脆弱生态系统的生物多样性。但在实际工作中,仍然需要加强脆弱生态系统的修复:一是对分布面积不太广泛的自然植被牛筋条-黄荆林和余甘子-明油子灌木林进行优先保护,从而避免因自然植被的单一化而导致的多样性大量丧失;二是对已有的草本层不发达的人工植被进行进一步修复。这需要决策者和林业工作者的共同努力,决策者应对破坏牛筋条-黄荆林和余甘子-明油子灌木林的开山修路、采石行为进行制止;林业工作者应以增加草本植物盖度为重点,保护和提高自然植被和人工植被的生物多样性。
柴宗新, 范建容. 2001. 金沙江干热河谷植被恢复的思考. 山地学报, 19(4): 381-384. DOI:10.3969/j.issn.1008-2786.2001.04.017 |
陈振耀, 梁铬球, 贾凤龙, 等. 2001. 广东南岭国家自然保护区大东山捕食性昆虫及其食性分析. 昆虫天敌, 23(1): 6-21. DOI:10.3969/j.issn.1674-0858.2001.01.002 |
丁建清, 付卫东. 1996. 生物防治——利用生物多样性保护生物多样性. 生物多样性, 4(4): 222-227. DOI:10.3321/j.issn:1005-0094.1996.04.007 |
侯美珍, 张永强, 王卫光. 2005. 玉米地捕食性节肢动物群落结构及动态分析. 中国生物防治, 21(4): 209-214. |
李昆, 罗长维, 陈友, 等. 2006. 元谋干热河谷生态恢复区昆虫多样性研究. 生态学杂志, 25(4): 417-422. DOI:10.3321/j.issn:1000-4890.2006.04.014 |
李巧, 陈又清, 陈祯, 等. 2006a. 元谋干热河谷直翅目多样性初步研究. 浙江林学院学报, 23(3): 316-322. |
李巧, 陈又清, 李从富, 等. 2006b. 元谋干热河谷象甲多样性初步研究. 西北林学院学报, 21(2): 102-106. |
李巧, 陈又清, 刘方炎, 等. 2007. 元谋干热河谷不同人工林中鞘翅目甲虫多样性比较. 生态学杂志, 26(1): 46-50. |
林荣华, 张润志, 田长彦, 等. 2003. 新疆棉田生态系统中捕食性节肢动物类天敌亚群落结构. 中国生物防治, 19(1): 1-5. |
罗淑萍, 张永强, 黄寿山. 2006. 不同抗性品种稻田捕食性节肢动物的群落结构. 昆虫知识, 43(4): 453-460. DOI:10.3969/j.issn.0452-8255.2006.04.005 |
马克平. 1994. 生物群落多样性的测度方法//中国科学院生物多样性委员会. 生物多样性研究的原理和方法. 北京: 中国科学技术出版社, 141-165.
|
任炳忠, 李典忠, 杨彦龙, 等. 2001. 吉林省农林天敌昆虫区系及多样性的研究(Ⅰ). 吉林农业大学学报, 23(4): 28-36. DOI:10.3969/j.issn.1000-5684.2001.04.008 |
师光禄, 王有年, 苗振旺, 等. 2006. 间种牧草枣林捕食性节肢动物群落结构的动态. 应用生态学报, 17(11): 2088-2092. DOI:10.3321/j.issn:1001-9332.2006.11.018 |
MoseleyR K, 唐亚. 2006. 云南干旱河谷150年来的植被变化研究及其对生态恢复的意义. 植物生态学报, 30(5): 713-722. DOI:10.3321/j.issn:1005-264X.2006.05.001 |
Colwell R K. 2005. Estimates: Statistical estimation of species richness and shared species from samples. Version 7. 5. User's Guide and application published at: http://purl.oclc.org/estimates.
|
Deborah L F, Robert F D. 2004. Predator diversity dampens trophic cascades. Nature, 429: 407-410. DOI:10.1038/nature02554 |
Hart D R. 2002. Intraguild predation, invertebrate predators, and trophic cascades in lake food webs. Journal of Theoretical Biology, 218: 111-128. DOI:10.1006/jtbi.2002.3053 |
Longcore T. 2003. Terrestrial arthropods as indicators of Ecological restoration success in coastal Sage scrub. Restoration Ecology, 11: 397-409. DOI:10.1046/j.1526-100X.2003.rec0221.x |
MacArthur R H. 1955. Fluctuations of animal populations and a measure of community stability. Ecology, 36: 533-536. DOI:10.2307/1929601 |
Odum E P. 1953. Fundamentals of Ecology. Philadelphia: Saunders, 132-157.
|
Tilman D, Lehman C, Bristow C E. 1998. Diversity-stability relationships statistical inevitability or ecological consequence. American Naturalist, 151: 277-282. |
Yachi S, Loreau M. 1999. Biodiversity and ecosystem functioning in a fluctuating environment:the insurance hypothesis. Proceedings of the National Academy of Sciences, USA, 96: 1463-1468. DOI:10.1073/pnas.96.4.1463 |