 2008, Vol. 44
2008, Vol. 44文章信息
- 陈晓鸣, 王自力, 陈勇, 赵杰军, 叶寿德, 王绍云.
- Chen Xiaoming, Wang Zili, Chen Yong, Zhao Jiejun, Ye Shoude, Wang Shaoyun
- 白蜡虫自然种群年龄特征生命表及主要死亡因素分析
- Age-Specific Life Table of Chinese White Wax Scale(Ericerus pela) Natural Population and Analysis of Death key Factors
- 林业科学, 2008, 44(9): 87-94.
- Scientia Silvae Sinicae, 2008, 44(9): 87-94.
-
文章历史
- 收稿日期:2006-12-05
-
作者相关文章


白蜡虫是一种重要的资源昆虫,其2龄雄若虫分泌的白蜡是一种重要的工业原料,具有重要的经济价值,广泛应用于化工、医药、食品、化妆品等行业。我国白蜡虫的培育和利用已有上千年的历史(王辅,1963a;周尧,1981;邹树文,1982),在云南、四川、湖南、贵州、陕西等省形成了昆虫产业。
白蜡虫1年发生1代,雌虫经1龄若虫、2龄若虫到成虫,为不完全变态;雄虫经1龄若虫、2龄若虫、前蛹、蛹到成虫,为完全变态。白蜡虫一生中在寄主植物上有2次爬行游动和转移,第1次爬行游动是卵孵化出1龄若虫在寄主上爬行,寻找适宜的叶片固定下来,生活一段时间,这段时间在白蜡生产上称为定叶;第2次转移发生在定杆期,1龄若虫脱皮后进入2龄,2龄若虫从叶片上转移到枝杆上生活,在白蜡生产上称为定杆。定杆后白蜡虫不再移动,雄虫交配后死亡,雌虫发育成熟产生下一代。白蜡虫生殖方式为卵胎生,种群发育整齐,无世代重叠。
前人在白蜡虫分类学(Chavannes,1819;Kuwana,1932)、生物学、生态适应性、泌蜡研究、泌蜡机理及生产技术(王辅,1963b;1986;柯志国,1981;吴次彬,1980;1989;吴次彬等,1987;1991;张长海,1991;张子有等,1985;1993;陈晓鸣等, 1997a;1997b;2007a;2007b;陈勇等,1998)等方面作了大量的工作。由于白蜡虫的种群数量大,取样和调查的难度很大,白蜡虫生命表的研究还未涉及,对白蜡虫自然种群的变动规律、繁殖能力、死亡因素等重大的生态学因素尚不十分清楚。生命表技术是从20世纪20年代提出的人口增长模型发展起来的,Morris等(1954)将生命表技术应用在昆虫自然研究上,此后,生命表技术应用广泛,并成为分析和研究昆虫种群动态的重要手段(丁岩钦,1994;徐汝梅等,2005)。研究白蜡虫自然种群生命表对揭示白蜡虫自然种群动态、数量变动规律、死亡因素、种群繁殖能力等方面具有十分重要的理论价值,对白蜡生产技术有重大的指导意义。本文通过对白蜡虫自然种群1995—2000年6个世代从卵、1龄若虫(定叶)、2龄若虫(定杆)、羽化、成虫和怀卵量等的系统研究和观察,组建了白蜡虫6个世代的平均生命表,分析了影响白蜡虫种群数量变动主要原因。
1 材料与方法 1.1 材料白蜡虫(Ericerus pela)采自云南昆明晋城白蜡园,寄主植物为女贞(Ligustrum lucidum)。生命表观察在昆明晋城白蜡园内进行。
1.2 方法 1.2.1 放养方式选择发育正常,无天敌危害的白蜡虫种虫(孕卵成熟的雌成虫),雌成虫孵化时放养在寄主植物枝条上,每头种虫放养1根枝条,枝条经修剪后,互不相交,每个枝条上只放养1头种虫,枝条基部涂凡士林,以防白蜡虫逃逸。放养每个世代白蜡虫50头,在发育的各个阶段采用生命表技术跟踪调查种群数量变动。
1.2.2 生命表调查生命表编制按照传统的年龄特征生命表方法(丁岩钦,1994;徐汝梅等,2005),具体方法如下。
1) 卵:白蜡虫为卵胎生,雌成虫发育成熟后,将卵产于虫囊中。随机抽取50头发育成熟的白蜡虫雌虫,解剖镜下实测白蜡虫产卵量。
2) 孵化率:随机抽取50头发育成熟的白蜡虫雌虫,在500 mL烧杯置于1头雌虫,用纱布扎紧烧杯口,置于20 ℃、70%相对湿度条件下孵化,统计孵化幼虫,采用孵化率=孵化若虫/怀卵量×100%计算孵化率。
3) 1龄若虫:1龄若虫孵化后经过爬行在寄主植物叶片上固定下来(定叶),白蜡虫雌虫在叶正面上定叶,雄虫在叶背面定叶。若虫定叶后,在蜕皮进入2龄若虫期前,按放养每头种虫的枝条调查每一片叶片上的若虫数量,实际统计出白蜡虫1龄若虫的存活数量,按存活率=1龄若虫存活数量/孵化若虫数量×100%计算1龄若虫存活率。
4) 2龄若虫:1龄若虫在叶片上蜕皮,蜕皮后2龄若虫从叶片上爬行到寄主植物枝条上固定下来终生不再移动(定杆),雄虫以群居方式定杆,定杆后很快分泌蜡丝,在雄虫化蛹前调查群体数量,调查方法为先测定白蜡虫群体定杆单位面积密度,然后调查白蜡虫泌蜡的面积,采用雄虫数=单位面积雄虫密度×泌蜡面积计算雄虫定杆数量;2龄雌虫在寄主植物的中上部枝条上定杆,散生分布,在2龄末期调查直接调查枝条上2龄雌若虫数量。
5) 蛹及羽化:白蜡虫雄虫为完全变态,经2龄若虫化蛹。按2龄雄若虫调查方法调查统计蛹数;随机抽样,采集白腊虫雄虫蛹的蜡块置于烧杯中用纱布扎紧烧杯口,在20℃、70%相对湿度条件下, 统计羽化和未羽化蛹,计算羽化率和成虫数量。
6) 成虫:白蜡虫雌成虫为不完全变态,2龄若虫蜕皮进入成虫期后,每月固定跟踪调查白蜡虫存活数量。雌成虫孕卵成熟后,随机抽取雌虫,剖开虫囊,将虫卵置于解剖镜下,根据雌雄卵具有不同颜色的特征,计数和统计性比。
7) 种群增殖能力:采用种群趋势指数I=N0(初始种群数量)/N1(下一代开始时种群数量)来描述白蜡虫种群增殖状况。
8) 样本数:在白蜡虫的各个龄期调查种群数量,每个世代调查枝条样本50个。生命表编制依据1995—2000年6个世代调查数据的平均数。
1.2.3 天敌及危害调查在白蜡虫发育的各个阶段调查白蜡虫天敌种类及危害,寄生蜂危害按采样的白蜡虫总数与被寄生数量统计寄生率;蜡象按抽样数量与危害数统计危害率;瓢虫对白蜡虫2龄雄若虫的危害,室内用白蜡虫2龄雄若虫饲养瓢虫,计算瓢虫对白蜡虫2龄雄若虫的取食量,蜡被受害状况按受害蜡被面/调查积蜡被面积×100%统计。寄生蜂种群动态调查采用黄色粘胶纸诱集小蜂成虫,在白蜡园内均匀挂置,每20天统计1次小蜂混合种群数量。
1.2.4 关键因子分析利用1995—2000年6个世代生命表数据,采用公式k=lglxi-lglxi+1计算出各龄期的K值,用致死力的分析方法作图,采用K值分析法中的K与ki回归系数分析法(Podoler et al., 1975)判断影响白蜡虫种群数量变动的关键因子。
2 结果与分析 2.1 自然种群生命表组建根据白蜡虫的生物学特征,从卵开始组建特定年龄生命表(表 1)。从白蜡虫生命表分析,白蜡虫自然种群在整个世代中,死亡率高达99%以上,其中,雌虫死亡率99%以上,雄虫死亡率96%以上。卵孵化占总死亡率的9.58%;若虫阶段在种群数量损失最大,阶段死亡率为85.25%,其中,1龄若虫阶段死亡率最高,为83.84%,达整个世代的70%左右,2龄幼虫的阶段死亡率45.50%,约占总死亡率的10%~15%;成虫期阶段死亡率为94.64%,占总死亡率的3%左右。高繁殖力(8 000~12 000头·♀-1)和高死亡率(>99%)是白蜡虫种群的显著特征,若虫2次爬行过程是种群数量损失的高峰期。
|
|

从白蜡虫自然种群存活状况来分析(图 1),白蜡虫自然种群具有繁殖率高,死亡率高,体型小等类似于r类有机体的特征。在生态适应性上,白蜡虫自然种群遵循r生存策略。白蜡虫一生中有2次若虫迁移,种群迁移能力较低,不能作长距离的迁移(一般在5 m以内),只能沿寄主枝条爬行寻找适合叶片定叶和枝条定杆,定杆后不能移动,后代保护机制较弱。白蜡虫以高繁殖率来补偿高死亡率,使种群得以生存和繁衍。

|
图 1 白蜡虫自然种群存活率 Figure 1 The livablity of Chinese white wax scale natural population |
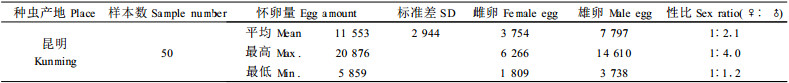
白蜡虫的怀卵量很大,昆明种虫平均怀卵量为11 553粒·♀-1,最高可达20 000粒·♀-1以上,最低4 000粒·♀-1,白蜡虫在卵成熟期就能从卵的颜色来判别雌雄,雌卵的颜色为深红色,雄卵颜色为黄色。测定表明,白蜡虫的雌雄性比一般在1:2左右,最高可达1:4,最低在1:1左右。雄虫比例高于雌虫(表 2)。调查发现,在白蜡虫雌成虫孕卵成熟期,除部分白蜡虫卵发育较差不孵化外,白蜡蚧长角象(Anthribus lajievorus)取食和小蜂寄生是影响白蜡虫孵化率的主要因素。在传统的白蜡虫种虫产区,白蜡蚧长角象取食卵和小蜂寄生卵,严重时受害率高达90%以上。
|
|
白蜡虫为卵胎生,雌成虫发育成熟后,虫体膨大,形成球囊状,将卵产于虫囊中,卵孵化后从虫囊中爬出。卵平均孵化率为90.42%,通常孵化率一般在85%~95%之间。死亡的主要原因是卵发育不全,未孵化。白蜡虫卵期天敌危害十分严重,常见的有白蜡蚧长角象和黑缘红瓢虫(Chilocorus rubidus)等,严重时,一头白蜡虫雌成虫囊中发现数十头白蜡蚧长角象取食虫卵,使白蜡虫种虫质量下降,对白蜡生产造成巨大的影响,严重时造成绝收。
2.2.3 若虫1) 1龄若虫:白蜡虫孵化后,1龄若虫在寄主植物枝条上爬行和游走,选择适合的叶片定叶。在定叶过程中,由于新孵若虫自身的保护机制差,1龄若虫数量从平均10 446头左右下降到平均2 849头,平均阶段死亡率为72.26%,占整个世代死亡率的60%左右。定叶过程是白蜡虫自然种群体数量损失最大的阶段,风、雨是若虫死亡的主要影响因子,风和雨将白蜡虫若虫中较弱的个体被淘汰,尤其是大风和暴雨将若虫吹走和冲刷,使白蜡虫种群数量受到很大的损失。
2) 2龄若虫:白蜡虫定叶大约1周后蜕皮进入2龄若虫,从叶片上迁移到寄主枝条上定杆。2龄若虫平均阶段死亡率为45.50%,约占总死亡率的10%~15%左右。2龄若虫死亡高峰发生在若虫蜕皮后定杆过程中,种群数量从平均2 849头下降到平均1 688头,大约40%的2龄若虫在迁移的过程中死亡。风、雨等气候因素是死亡的主要原因。定杆后,2龄雄若虫种群数量从平均1 688头下降到化蛹时平均771头,平均阶段死亡率为54.32%;2龄雌若虫数量从平均327头下降到进入成虫初期平均149头,死亡率为54.43%。定杆后白蜡虫2龄雄若虫开始分泌白蜡覆盖和保护虫体,雌虫体壁逐渐角质化,形成较完善的保护机制。寄生性和捕食性天敌成为主要死亡原因。
2.2.4 蛹及雄成虫白蜡虫雄虫为完全变态,平均771头2龄若虫化蛹,孵化为成虫448头,有一半以上的蛹可以羽化为成虫,平均孵化率为63.29%,阶段死亡率为41.89%,占总死亡率的2.80%。雄成虫在7—8月羽化,羽化后就近寻找雌成虫交配,交配后死亡。一头雄成虫可以与多头雌成虫交配,雄成虫可以存活8~10 d。
2.2.5 雌成虫进入成虫后,雄成虫交配后自然死亡,雌成虫则要经历长达近8~9个月(9月—翌年5月)的生长发育,孕卵,产生后代。在成虫初期,种群数量在平均597头,到成虫末期时只剩下平均32头,雌成虫阶段的平均死亡率在94.64%,占整个世代死亡率的4.89%。雌成虫初期死亡率较高,11月前的死亡率占整个雌成虫期的80%以上,11月后,雌成虫死亡较少,种群数量变动较为平稳(图 2),这与8—11月的小蜂高发时期相吻合(图 3)。白蜡虫雌成虫主要死亡原因是小蜂寄生、蜡象和瓢虫取食等天敌危害。

|
图 2 白蜡虫雌成虫存活曲线 Figure 2 The surival curve of female adult of Chinese white wax scale |

|
图 3 寄生蜂混合种群的数量动态曲线 Figure 3 The mix populations dynamic of parasitic wasps |
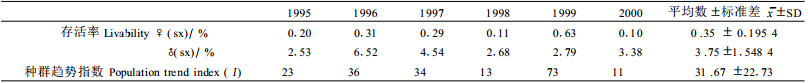
从白蜡虫自然种群的存活率和种群趋势指数来分析(表 3),种群最低存活率为0.10%,最高存活率为0.63%,雌虫死亡率达99%以上,雄虫死亡率在96%左右(扣除交配后的自然死亡)。白蜡虫的卵量大,但种群的死亡率也很高,不到1%的雌成虫存活下来。调查结果表明(表 3),种群趋势指数平均为32,最高为73,最低为11,在每个时代调查的50个样本中,最高种群趋势指数可达100~200,最高繁殖倍数是最低繁殖倍数的10~15倍,甚至更高。说明白蜡虫的种群增殖能力与环境因素密切相关,在适宜的环境条件下,白蜡虫种群具有较大的增殖空间。
|
|
白蜡虫若虫的死亡高峰发生在定叶和定杆的过程中,70%~80%的若虫在2次爬行和迁移中死亡。由于白蜡虫若虫的保护机制较弱,大风和大雨会导致若虫大量死亡。试验证明(张子有等,1985),新孵化的1龄若虫在风速4 m·s-1以上,50%以上的若虫被吹走,2龄若虫的抗风力大于1龄若虫。在昆明地区,白蜡虫定杆过程中风较大,观察发现,体弱的白蜡虫个体在爬行和转杆的过程中被风吹走。雨水对白蜡虫死亡的影响主要表现在几个方面,大雨会注入白蜡虫虫囊中,雨水浸泡虫卵,使卵不能孵化而死亡;在若虫定叶、定杆转移过程中,雨水冲刷,特别是大暴雨,会使白蜡虫若虫有较大的损失,而且叶面部分积水,也会使定叶的白蜡虫若虫死亡;白蜡虫进入成虫后,长时期的高湿度对白蜡虫的存活不利,导致死亡率较高,种群不能正常延续(陈晓鸣等,2007a;2007b),岑明等(1993)在广西的调查也说明雨量较大的地区不适于白蜡虫生存。
2.3.2 天敌白蜡虫一生都受到天敌的危害,在若虫期,瓢虫类天敌取食若虫;2龄若虫定杆后,雄虫开始泌蜡,瓢虫破坏蜡被,取食2龄雄若虫,寄生蜂对白蜡虫2龄若虫危害十分严重,多种寄生蜂寄生雌雄若虫;在成虫阶段,白蜡虫死亡的主要原因是瓢虫取食、小蜂寄生、蜡象取食卵等。通过对白蜡虫产区调查,主要天敌有16种,分属于膜翅目和鞘翅目(表 4)。
|
|

1) 寄生类天敌:白蜡虫的寄生性天敌主要是小蜂类,白蜡虫寄生蜂有20多种,常见的有11种,白蜡虫花翅跳小蜂主要寄生白蜡虫雄虫, 中华花翅跳小蜂主要寄生白蜡虫卵,白蜡虫花翅跳小蜂、白蜡虫阔柄跳小蜂、白蜡虫蓝绿跳小蜂、白蜡虫啮小蜂、白蜡虫纹翅跳小蜂、日本食蚧蚜小蜂、黑褐纹翅跳小蜂等寄生于白蜡虫的幼虫和成虫体内。
在昆明白蜡园内,中华花翅跳小蜂对卵囊(孕卵成熟的雌成虫)平均寄生率为35%,平均每虫囊寄生1.2头,最高可达27头。1999—2000年在昆明白蜡园内诱集到3 219头小蜂,其中白蜡虫花翅跳小蜂占45.90%,白蜡虫阔柄跳小蜂占37.36%,中华花翅跳小蜂占11.39%,黑褐纹翅跳小蜂占1.96%,白蜡虫啮小蜂占1.03%,其余6种蜂仅占2.36%。白蜡虫花翅跳小蜂、白蜡虫阔柄跳小蜂和中华花翅跳小蜂为优势种群。
在昆明地区,寄生蜂种群呈现出3个高峰期,分别发生在8—9月、11—12月和4—5月(图 3),4—5月是白蜡虫卵发育成熟期,主要是中华花翅跳小蜂寄生;8—9月是白蜡虫泌蜡后期,主要是白蜡虫花翅跳小蜂、白蜡虫阔柄跳小蜂和中华花翅跳小蜂;11—12月是成虫初期,优势寄生蜂为白蜡虫花翅跳小蜂和中华花翅跳小蜂, 优势种群的寄生率都在30%以上(表 5)。
|
|

2) 捕食类天敌:捕食白蜡虫的主要天敌是蜡象和瓢虫,危害白蜡虫各个虫态。常见的主要种类有白蜡虫长角象(蜡象)、红点唇瓢虫、黑缘红瓢虫、二双斑唇瓢虫、异色瓢虫和七星瓢虫。蜡象主要取食白蜡虫卵,瓢虫取食白蜡虫各个虫态的组织。根据对云南、四川等地产虫区调查,种虫老产区蜡象危害率较高,四川金口河、四川喜德的危害率在90%以上,云南昭通的危害率在60%左右,每头种虫中最少的有3头,最多的有48头蜡象,严重地影响白蜡虫种虫的质量和产量。瓢虫对白蜡生产的危害主要是咬开蜡被取食白蜡虫幼虫,造成白蜡生产损失。黑缘红瓢虫、二双斑唇瓢虫是白蜡虫2龄雄幼虫主要天敌,1头黑缘红瓢虫一生可以取食1 000头以上的白蜡虫2龄雄若虫,损坏蜡被12~15 cm2, 二双斑唇瓢虫1头虫一生取食600头以上的白蜡虫2龄雄若虫,损坏蜡被8~10 cm2, 2种瓢虫使白蜡产量损失20%以上(表 6)。
|
|

根据白蜡虫的生物学特征和生命表调查数据,用致死力的分析方法作图(图 4),对影响白蜡虫种群动态的关键因子进行分析。从图 4中可以看出,k2(1龄若虫死亡)和k6(雌成虫死亡)数量变动对K(世代总致死力)的波动影响较大,是影响全年种群动态变动的关键因素。按照Podoler等(1975)判断生命表关键因子的方法,采用回归方法对k2和k6与K进行回归,得到:K=0.109+3.368k2;K=2.190+0.755k6。根据这个判断方法,回归方程中回归系数b值大的为关键因子,从2个回归方程中b值比较得出,k2的b值为3.368>k6的b值(0.755),所以k2是影响种群动态的关键因素。进一步分析k2和k6变动的原因,1龄若虫大量失踪主要是风、雨等气候因子导致,成虫死亡原因主要是天敌危害所致。分析表明,风雨对白蜡虫种群变动的影响大于天敌危害。

|
图 4 白蜡虫自然种群动态关键因素分析 Figure 4 The key factor analysis of natural population dynamic of Chinese white wax scale |
本研究揭示了白蜡虫自然种群特征和动态规律,白蜡虫具有高繁殖力、高死亡率的种的显著特征。由于若虫保护机制较弱,种群逃避风险能力较差,若虫期是种群死亡率最高的时期,主要死亡原因是气候因子;成虫期是阶段死亡率最高的时期,主要死亡原因是天敌;在无灾变性气候和天敌严重的条件下,白蜡虫种群的世代趋向指数在30以上。白蜡虫以高繁殖力来补偿高死亡率,使种群生存和延续。
若虫2次迁移是白蜡虫种的特征,白蜡虫种群一生中的2次迁移过程是白蜡虫种群自身保护机制最薄弱的时期,也是白蜡虫死亡最高的阶段,这个阶段的死亡率占种群总的死亡率的80%以上。一般认为(张长海等,1999)白蜡虫死亡的主要原因是天敌,实际上,白蜡虫在2次迁移中的损失对白蜡虫种群影响最大,是白蜡虫生产的关键时期,风、雨等气象因子是白蜡虫死亡的关键因素。在2次迁移中,风、雨淘汰了白蜡虫自然种群中体质较弱的个体。80%以上的若虫死亡,这是由于白蜡虫种的遗传特征所决定的,即便在无大风、大雨的理想状况下,在定叶过程中,也有相当数量1龄若虫在爬行过程中死亡,不能定叶。
白蜡虫定杆后,雄若虫分泌白蜡被覆盖虫体,雌虫进入成虫后,体壁逐渐角质化,形成较完善的保护机制,风雨等气候因子对白蜡虫的影响降低,寄生性和捕食性天敌成为白蜡虫死亡的重要原因。白蜡虫雄虫定杆后,在适宜的泌蜡区域,影响白蜡生产的主要原因是天敌,寄生和捕食害率可高达40%以上,控制天敌是提高白蜡产量的重要途径。此外,白蜡虫雄虫定杆后,还受到密度制约的影响,白蜡虫雄虫以群居方式定杆,定杆初始密度为110~130头·cm-2,到羽化前密度为80~90头·cm-2,在没有天敌的情况下,随着若虫体增长,由于定杆后若虫不能迁移,为了争夺有限的空间和食物资源,竞争力较弱的个体在竞争中死亡。
白蜡虫雌成虫阶段的死亡死亡率为95%以上,死亡主要原因是天敌,天敌危害率在30%~60%,高可达90%以上。白蜡虫的种群时代倾向值在10~40之间,最高的繁殖倍数可到100倍以上,说明雌成虫具有较大的增殖空间,有效地控制天敌可以使白蜡虫自然种群数量增值,从而获得种虫高产。除天敌危害外,雌成虫的生长发育同样存在密度制约效应,白蜡虫雌虫定杆时,虫体直径只有约0.1 cm大小,到发育成熟时,虫体直径在0.5~1.2 cm之间,由于不能迁移,如果定杆密度过大,会造成雌成虫为争夺有限的空间和食物资源而导致大量死亡或发育为相互挤压,畸形的种虫,使种虫质量下降,同样影响白蜡生产。因此,合理的放养密度也是提高白蜡虫种虫产量和质量的重要措施。
岑明, 计鸿贤, 林发源. 1993. 人降雨量对广西白蜡虫死亡及其种虫、白蜡产量的影响. 林业科学研究, 6(2): 194-199. |
陈晓鸣, 陈勇, 叶寿德, 等. 1997a. 白蜡虫孵化行为研究. 林业科学研究, 10(2): 149-153. |
陈晓鸣, 陈勇, 叶寿德, 等. 1997b. 白蜡虫在寄主植物上的分布特征研究. 林业科学研究, 10(4): 415-419. |
陈晓鸣, 王自力, 周朝鸿, 等. 1998. 白蜡虫泌蜡研究Ⅰ.不同地理种源泌蜡比较. 林业科学研究, 11(1): 35-38. |
陈勇, 陈晓鸣, 王自力, 等. 1998. 白蜡虫泌蜡研究Ⅱ.不同寄主植物上泌蜡比较. 林业科学研究, 11(3): 285-288. DOI:10.3321/j.issn:1001-1498.1998.03.010 |
陈晓鸣, 王自力, 陈勇, 等. 2007a. 影响白蜡虫泌蜡主要气候因子及白蜡虫生态适应性分析. 昆虫学报, 50(2): 1136-1143. |
陈晓鸣, 王自力, 陈勇, 等. 2007b. 环境因子对白蜡虫泌蜡的影响. 生态学报, 27(1): 103-119. |
丁岩钦. 1994. 昆虫数学生态学. 北京: 科学出版社, 153-214.
|
柯治国. 1981. 白蜡虫的分布与生态因子的分析. 昆虫知识, 18(6): 257-259. |
王辅. 1963a. 我国古代繁殖白蜡虫的记载. 昆虫知识, 9(9): 178. |
王辅. 1963b. 白蜡虫雌雄群体的生态适应性及其在生产上的应用. 林业科学, 8(2): 171-175. |
王辅. 1986. 发展白蜡生产的途径问题——兼与吴次彬同志商榷. 动物世界, 3(2-3): 73-84. |
吴次彬. 1980. 对"白蜡虫雌雄群体的生态适应性及其在生产上的应用"一文商榷. 林业科学, 16(4): 296-301. |
吴次彬, 王一丁, 林一中, 等. 1987. 白蜡虫几个地方种虫经济性状的比较研究. 动物学研究, 8(3): 322-327. |
吴次彬. 1989. 白蜡虫及白蜡生产. 北京: 中国林业出版社, 1-115.
|
吴次彬, 朱伟, 冉江红. 1991. 对白蜡虫种虫和白蜡生产布局问题的探讨. 四川大学学报:自然科学版, 28(3): 345-352. |
张长海. 1991. 白蜡虫在我国的地理分布. 林业科学研究, 4(2): 192-195. |
张长海, 刘化琴, 李力, 等. 1999. 白蜡虫种虫生产中三红三黑原因研究. 林业科学研究, 12(1): 37-41. DOI:10.3321/j.issn:1001-1498.1999.01.007 |
张子有, 邵孟明, 马继明, 等. 1985. 不同生态环境下繁育白蜡虫(Ericerus pela Chavannes)研究. 动物世界, 2(3-4): 227-232. |
张子有, 齐胜利, 张良玉. 1993. 白蜡虫生活在不同生态环境下繁殖力观察研究. 昆虫知识, 30(5): 297-299. |
邹树文. 1982. 中国昆虫学史. 北京: 科学出版社, 112-117.
|
周尧. 1981. 中国昆虫学史. 西安: 昆虫分类学报社, 41-42.
|
徐汝梅, 成新跃. 2005. 昆虫种群生态学. 北京: 科学出版社, 38-64.
|
Chavannes M A. 1819. Notice Sur deux Coccus ceriferes du Bresil. Annual Society Entomology France, 6(2): 139-145. |
Kuwana I. 1932. The Chinese white wax scale. Philippine Journal of Science, 22(4): 393-405. |
Morris R F, Miller C A. 1954. The development of life tables for the spruce budworm. Canada Journal of Zoology, 32: 283-301. |
Podoler H, Roglers D. 1975. A new method for the identification of key factor from life table data. Journal of Animal Ecology, 44: 85-114. |