 2008, Vol. 44
2008, Vol. 44文章信息
- 汤景明, 翟明普, 崔鸿侠.
- Tang Jingming, Zhai Mingpu, Cui Hongxia
- 壳斗科三树种幼苗对不同光环境的形态响应与适应
- Morphological Responses and Adaptation of Seedlings of Three Tree Species of Fagaceae Family to Different Light Environments
- 林业科学, 2008, 44(9): 41-47.
- Scientia Silvae Sinicae, 2008, 44(9): 41-47.
-
文章历史
- 收稿日期:2007-09-18
-
作者相关文章


2. 北京林业大学省部共建森林培育与保护教育部重点实验室 北京 100083
2. Key Laboratory of Forestry Cultivation and Conservation of Education Ministry, Beijing Forestry University Beijing 100083
幼苗作为森林植物个体发育中的一个重要阶段,是森林植物生活史中最脆弱、最敏感的时期(李俊清等,2001;韩有志等,2002)。幼苗的生长受自身遗传物质和环境因素的综合影响,是对环境不断适应的过程。在此过程中,受到光照、水分、温度等生态因子及其相互作用的影响,其成活、生长发育状况直接关系到种群的生存和发展,以及森林的发育、演替和更新。在自然界中,生长在不同环境中的同种植物,常在形态、生理、生物量等方面产生可塑性变化以适应变化的环境(王艳红等,2005)。
光照是植物生长发育过程中的重要环境因子之一,影响植物生长环境、光合作用和形态结构。植物对光环境的反应受当前光环境和其历史光环境的复合影响。目前,国内外关于植物对光环境的响应与适应性的研究主要集中在相同环境下不同物种对光的反应、在遮荫下生长的植物移植到强光环境下其发育表型的短期响应以及在不同强度的遮荫对各项生态指标的影响等方面(郭志华等,2003;陈圣宾等,2005)。以往工作重点研究不同光环境对植物形态、叶片解剖结构、生长发育、生物量分配及生理特性方面的影响,揭示了不同植物对光环境的适应策略(肖春旺等,1999;孙书存等,1999;温达志等,1999;Agyeman et al., 1999;徐程扬,2001;Sack et al., 2002;何维明等,2002;郭志华等,2003;王艳红等,2005;安锋等,2005;冉春燕等,2006)。
锥栗(Castanea henryi)、水青冈(Fagus longiptiolata)和青冈(Cycloblanopsis glauca)是我国常绿落叶阔叶混交林的主要组成树种,其幼苗的生长对森林的发育、演替和更新具有重要意义。本研究以人工控光条件下锥栗、水青冈和青冈1年生幼苗为对象,研究其对不同光环境的形态响应与生态适应性,进一步揭示山地常绿落叶阔叶混交林主要树种更新机制,为我国南方常绿落叶阔叶混交林快速恢复、人工育苗、造林技术提供基础资料和理论依据。
1 试验地概况与研究方法 1.1 试验地概况试验地位于湖北省鹤峰县木林子自然保护区内(30°27′ N,110°23′ E,海拔1 280 m)。属中亚热带湿润季风气候。年平均气温11.9 ℃。1月平均气温9.7 ℃,极端最低气温-17.1 ℃。7月平均气温28.6 ℃,极端最高气温32 ℃。年降水量1 800 mm,相对湿度85%,无霜期260 d。土壤以黄棕壤和棕壤为主,pH值4.5~6.5。自然植被主要为山地常绿落叶阔叶混交林。
1.2 研究方法 1.2.1 人工控光育苗试验在木林子自然保护区空旷苗圃地进行。光环境控制采用人工模拟试验法,用黑色聚乙烯遮荫网遮荫,并分为3个光梯度,即全光(记为L0)、一层网遮荫(记为L1)、二层网遮荫(记为L2),其中L1和L2相当于全光环境下光照强度的56%和32%。每个光处理面积7 m2,分3个小区,每小区面积2.25 m2。2004年11月,取林中A层土壤并混合均匀后铺于苗床中,在每个光处理的3个小区分别播种锥栗、水青冈、青冈种子100粒,播种后覆土2 cm。常规方法育苗。
1.2.2 测定方法2005年11月上旬,在苗木停止生长但尚未落叶时,在每小区中央选30株幼苗全部挖出,逐株测定苗高(从地面到苗木顶芽的高度)、地径(苗木土痕处的粗度)、主根长、叶片数、叶长(1 cm以上叶片的长度)和最大叶宽。随机选取大小不等的60片叶子,实测每片叶的叶面积。再根据叶片长和最大宽度,用回归法求得:锥栗单叶面积=0.499+(叶长×叶最大宽)×0.631(R2=0.960,P<0.01,n=60);水青冈单叶面积=0.468+(叶长×叶最大宽)×0.583(R2=0.981,P<0.01,n=60);青冈单叶面积=0.188+(叶长×叶最大宽)×0.577(R2=0.980,P<0.01,n=60)。根据所测每片叶的叶长和叶宽计算每片叶的单叶面积,据此分别求出3种光照强度下锥栗、水青冈、青冈幼苗的叶面积。把着生于主茎上的枝条作为一级侧枝,统计一级侧枝的总数和有侧枝的幼苗株数,并计算有侧枝率。随机选取其中的10株幼苗,分叶片、侧枝、主茎、侧根(含须根)、主根称鲜质量,称重后将样品在烘箱中65 ℃烘12~24 h后称其干质量。在此基础上,计算比叶面积(单株叶面积与叶干质量之比)、叶面积比(单株叶面积与总干质量之比)、高径比(苗高与地径之比)和茎根比(地上生物量与根生物量之比)。苗木质量指数(quality index; QI)按下式进行计算:QI=苗木总干质量/[(苗高/地径)+(茎干质量/根干质量)]。式中,苗高/地径是苗高以cm、地径以mm为单位的高径比。
1.3 统计分析方法利用SPSS软件进行方差分析,用Ducan法进行多重比较,并用字母法标记。
2 结果与分析 2.1 不同光环境下幼苗的形态特征从表 1可知,全光(L0)下锥栗和水青冈的苗高、地径均显著高于L1、L2光强下的苗高、地径,L1和L2光强下锥栗和水青冈的苗高、地径没有明显差异。青冈苗高、地径在L0、L1、L2 3种光强下均无显著差异。光照环境对3个树种幼苗主根的生长均有明显影响,随着光照强度降低,幼苗主根生长量呈下降趋势。这表明,光照能促进地下根系的生长,当光照强度减弱时,主根深度相应变浅。这是因为在全光环境下,土壤表层土受较强的太阳光直射、风吹等原因,蒸发量较大,含水率较低,播种深度仅为2 cm的坚果萌发生根后,为了维持出土子叶的水分平衡,在这种水分胁迫下幼苗通过主根的伸长深入下层土壤中吸收水分;同时植株通过较发达的主根来适应较强光照条件下幼苗的快速生长,并满足对土壤水分和养分的需求。由于遮荫处理下幼苗生长的环境空气湿度大,土壤含水量高,叶片蒸腾作用弱,加上与之竞争的杂草少且生长较弱,幼苗不需要向根系投入更多的生物量即可获得生长所需水分和养分。
|
|
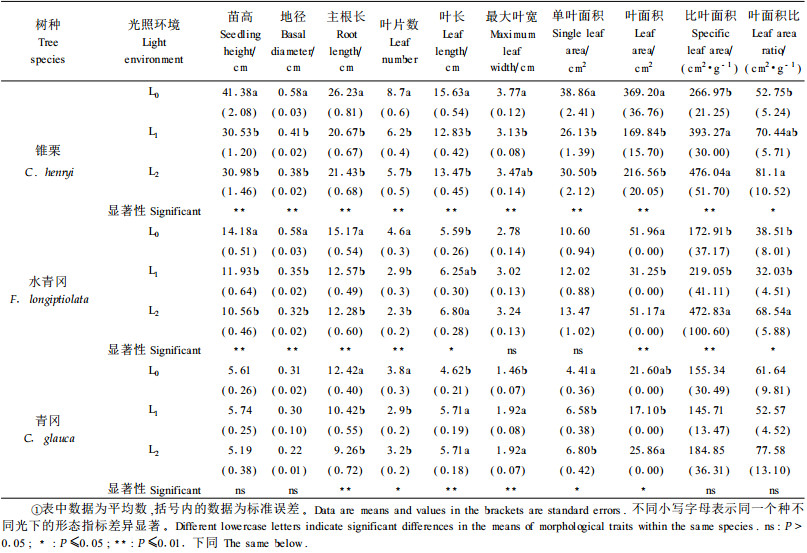
随着光照强度的减弱,3树种幼苗的现存叶片数呈明显减少趋势。青冈幼苗个体小,叶数量少,这一规律表现得不如锥栗和水青冈明显。随着光照减弱,锥栗幼苗的叶长、叶宽、单叶面积呈明显减少的趋势,而水青冈和青冈则相反,但水青冈叶宽和单叶面积未达显著性差异。这说明,锥栗更适应于在全光下生长,植株整体生长均较遮荫条件下好,相应地其叶片也得到正常的生长发育。3树种幼苗的叶面积随着光照的变化也发生相应的改变,并表现出明显差异。锥栗叶面积在全光(L0)最大,水青冈和青冈幼苗叶面积均在一层网遮荫(L1)最小。3树种幼苗的比叶面积和叶面积比均不同程度地受光环境的影响,并表现出随光照减弱而增大的趋势。锥栗、水青冈幼苗的比叶面积随光照减弱逐渐增大,全光(L0)下的比叶面积明显低于L1、L2光环境,L1和L2光强下的比叶面积无明显差异。青冈幼苗的比叶面积随光照减弱呈增加趋势,但L0、L1和L2 3种光环境下的比叶面积无明显差异。3树种幼苗的叶面积比表现出随光照减弱呈增加趋势,锥栗幼苗的叶面积比的这一变化规律比水青冈、青冈幼苗明显。叶面积比的这种反应,表明幼苗在遮荫条件下的叶片比全光下的薄而宽大,以适应弱光下捕获光能的需要。因为比叶面积随光照减弱而增加,有助于提高单位叶生物量的表面积,增大叶截取光资源的有效面积,提高光能利用效率,以适应较弱的光环境(Pearcy,1999)。
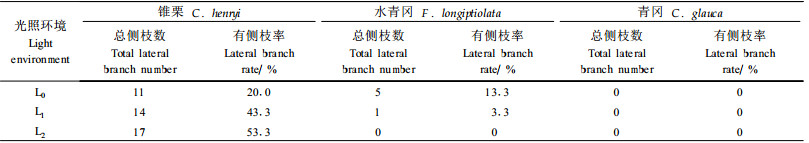
从表 2可以看出,随着光照的减弱,锥栗幼苗的侧枝率逐渐增加,L0、L1、L23种光强下分别为20.0%、43.3%、53.3%;水青冈侧枝率逐渐减少,分别为13.3%、3.3%、0;青冈在3种光强下均没有分枝。这说明,光照强度对幼苗分枝具有一定的影响,树种不同,其幼苗的分枝对光环境的反应存在较大的差异。研究表明,遮荫条件下,植株采取加强顶端优势的对策,限制侧枝的发育,使侧枝数会比全光下显著减少。通常具有较高分枝率的枝系统有利于强光下树冠对光辐射的截获,而低分枝率的枝系统则可以使树木叶片在弱光下排列更有效(安锋等,2005;Hart,1998;Canham,1998;Gilmore et al., 1997;徐程扬,2001)。本研究中,1年生水青冈幼苗侧枝对光环境的反应支持了这一结论。
|
|
从表 3可以看出,不同树种、不同光强下的幼苗生物量有明显差异。遮荫条件下生长的锥栗、水青冈幼苗单株生物量比全光低,其中锥栗减少幅度最大,L1和L2的干物质量仅为L0的32.6%和38.5%,水青冈次之,相应的比例为67.5%和53.4%,但锥栗、水青冈单株生物量在L1和L2之间没有显著差异。青冈幼苗单株生物量在各光环境下非常接近,L0略高于L1和L2。光照强度还显著影响了各器官干物质的积累。两遮荫条件下(L1和L2)的锥栗、水青冈,其叶、主茎、主根、地上、地下干重均低于全光条件,且锥栗的变化幅度大于水青冈,但L1与L2之间差异不大。不同光照条件下青冈幼苗各器官生物量干重仅在数量上略有差异,但没有达到显著性水平。很多研究都发现:在强光照射下,由于蒸腾作用增强,幼苗增加根部的投资,将生物量相对多地分配到地下根系,来解决对水分和矿质养分的需求,在弱光照条件下,幼苗将更多的生物量分配到地上部分特别是叶器官上,来增加对光能的捕获,提高光合作用能力(Poorter,1999;King,2003;陈圣宾等,2005)。在中等遮荫下,地上地下生物量平衡,生物量积累达到最大(Gardiner et al., 1998)。本研究并没有发现3树种幼苗在不同的光照强度下生物量分配格局有明显变化,这可能是因为本研究对象为3树种的1年生播种苗,幼苗个体小,本身固定的有机物有限,幼苗对光强的可塑性不如成熟个体(Rosenthal et al., 1994),使得幼苗的生物量分配没有表现出随光强变化而变化。
|
|

运用非线性幂函数模型W=a(D2H)b进行拟合,建立人工控光不同光照强度下锥栗、水青冈和青冈幼苗生物量(W)与苗高(H)和地径(D)的9个回归模型,其参数a、b及相关性指标R2见表 4。
|
|
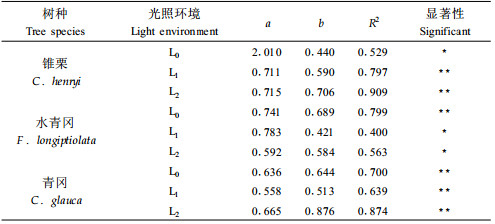
从表 4看出,各模型拟合效果好,均达到5%或1%的显著性水平。不同光环境下锥栗、水青冈和青冈幼苗生物量与苗高、地径的变化趋势遵从幂函数模型。回归模型中b值表示幼苗生物量的增加速率,即幼苗D2H每增加1 cm3时,幼苗生物量增加的速率。不同光强下,锥栗幼苗D2H值每增加1 cm3时,生物量将增加0.440~0.706 g;水青冈幼苗D2H值每增加1 cm3时,生物量将增加0.421~0.689 g;青冈幼苗D2H值每增加1 cm3时,生物量将增加0.513~0.876 g。回归模型中a值表示幼苗开始积累生物量时D2H的理论值。锥栗、水青冈和青冈幼苗在3种光强下的a值分别在0.711~2.010 cm3、0.592~0.783 cm3和0.558~0.665 cm3之间变化,当幼苗D2H分别达到这3个变动范围内时,锥栗、水青冈和青冈幼苗才开始生物量积累。
2.3 不同光环境幼苗质量评价从表 5中可以看出,不同光环境下锥栗高径比表现为L0<L1<L2,但没有显著性差异。水青冈、青冈高径比均表现为L0<L2<L1,并有显著性差异,其中水青冈L0与L1、L2有显著差异,L1和L2之间没有显著差异,青冈L0、L1和L2之间有显著差异。不同光环境下锥栗幼苗茎根比表现为L0<L2<L1,水青冈L2<L1<L0,青冈L1<L0<L2,但均没有显著差异。3树种幼苗在全光下质量指数最高,锥栗、水青冈幼苗在全光下质量指数显著高于2个遮荫处理(L1和L2),但L1和L2之间的差异没有显著性水平。青冈幼苗在全光下质量指数略高于2个遮荫处理(L1和L2),在3种光环境下均没有显著性差异。
|
|

锥栗、水青冈和青冈1年生幼苗对不同光环境产生不同程度的形态响应。幼苗的生长指标随光环境的变化程度表现出锥栗>水青冈>青冈。随着光照的减弱,3树种幼苗的苗高、地径、主根和生物量等生长指标呈下降趋势。锥栗幼苗的叶片数、叶长、叶宽、单叶面积随光照减弱逐渐减少,水青冈和青冈幼苗的叶片数随光照减弱呈下降趋势,而其叶长、叶宽、单叶面积则呈明显增加趋势。锥栗在全光照环境幼苗叶面积最大,而水青冈和青冈幼苗叶面积均以56%光照强度最小,其原因有待于进一步研究。3树种幼苗比叶面积和叶面积比随光照减弱呈增加趋势。光照对不同树种幼苗的分枝具有不同的影响。随着光照的减弱,锥栗幼苗的分枝率逐渐增加,水青冈幼苗的分枝率逐渐减少,青冈幼苗尚未产生分枝。木本植物幼苗的分枝程度受环境信号、发育阶段和遗传因素的综合调控,植物激素参与这些信号的整合(巩鹏涛等,2005;王冰等,2006)。本研究中3种壳斗科树种的1年生幼苗在不同光环境下分枝的差异涉及到十分复杂的植物激素调控和信号转导途径,并与光环境相关联,其作用机理有待于进一步深入研究。
植物形态上的差异是环境资源的有效供给能力和植物体对资源利用能力的综合表现。环境条件的变化,引起植物形态结构和生物量的变化,形态结构的变化又会改变植物所处的微环境。人工控光环境下更新树种幼苗的形态可塑性响应差异,反应了更新树种不同的生态对策和适应性。本研究发现,锥栗1年生幼苗在全光照条件下生长量最大,在不同程度的遮荫环境下生长受到明显抑制。野外调查还发现,锥栗在阳坡或开阔地生长良好,在郁闭的天然林林冠下未见锥栗更新苗,这说明锥栗为喜光阳性树种。本研究水青冈1年生幼苗在全光照条件下生长较遮荫条件下好,表明水青冈在更新过程中,在幼苗定居阶段需要一个较好的光照条件,但较高的幼树能在林下荫蔽的条件下持续生存很长的时间(Peters,1997;郭柯,2003)。可见,水青冈为后期耐荫种。本研究中青冈1年生幼苗对光环境的改变反应不敏感,而且野外调查也发现,在较郁闭的林下有生长良好的更新苗,这说明青冈为适应能力强的广适种。
3树种幼苗均表现出在全光下质量指数最高,锥栗、水青冈幼苗明显大于遮荫处理,而青冈幼苗差异不大。因此可选择开阔苗圃地,在全光照环境下培育锥栗、水青冈和青冈优质实生苗。由于质量指数综合了幼苗苗高、地径等主要形态因子和生物量指标,可以反映苗木各部分之间的协调和平衡,克服单个形态因子评价苗木质量的不足,可为判断更新苗移植成活率提供依据(沈国舫,2001)。锥栗、水青冈和青冈是鄂西南山地常绿落叶阔叶混交林的主要建群种,在常绿落叶阔叶混交林恢复与演替过程中起着重要的作用。在常绿落叶阔叶混交林恢复过程中,要根据3树种幼苗对环境胁迫的响应与适应,因地因林制宜,制定科学的恢复策略。在皆伐迹地,选择锥栗作为造林先锋树种,实现森林快速更新;在择伐地或出现林窗干扰的天然林中,选择水青冈和青冈作为补植(补播)树种,促进群落进展演替。在人工混交林营造和人工促进天然更新等营林实践中,选择青冈作为混交树种和林下引进树种。
安锋, 林位夫. 2005. 植物耐荫性研究的意义与现状. 热带农业科学, 25(2): 68-74. DOI:10.3969/j.issn.1009-2196.2005.02.016 |
陈圣宾, 宋爱琴, 李振基. 2005. 森林幼苗更新对光环境异质性的响应研究进展. 应用生态学报, 16(2): 365-370. DOI:10.3321/j.issn:1001-9332.2005.02.034 |
巩鹏涛, 李迪. 2005. 植物分枝发育的遗传控制. 分子植物育种, 3(2): 151-162. DOI:10.3969/j.issn.1672-416X.2005.02.001 |
郭柯. 2003. 山地落叶阔叶林优势树种米心水青冈幼苗的定居. 应用生态学报, 14(2): 161-164. DOI:10.3321/j.issn:1001-9332.2003.02.001 |
郭志华, 臧润国, 奇文清, 等. 2003. 中亚热带2种原始兰科植物对变化光环境的响应与适应. 林业科学, 39(3): 23-29. |
韩有志, 王政权. 2002. 森林更新与空间异质性. 应用生态学报, 13(5): 615-619. DOI:10.3321/j.issn:1001-9332.2002.05.024 |
何维明, 董鸣. 2002. 异质光环境中旱柳的光截取和利用反应. 林业科学, 38(3): 7-13. DOI:10.3321/j.issn:1001-7488.2002.03.002 |
李俊清, 臧润国, 蒋有绪. 2001. 欧洲水青冈构筑型与形态多样性研究. 生态学报, 21(1): 151-155. |
冉春燕, 陶建平, 宋利霞. 2006. 喜树幼苗形态和光合作用对不同光环境的适应. 西南师范大学学报:自然科学版, 31(2): 142-146. |
沈国舫. 2001. 森林培育学. 北京: 中国林业出版社.
|
孙书存, 陈灵芝. 1999. 不同生境中辽东栎的构型差异. 生态学报, 19(3): 359-364. DOI:10.3321/j.issn:1000-0933.1999.03.012 |
王冰, 李家洋, 王永红. 2006. 生长素调控植物株型形成的研究进展. 植物学通报, 23(5): 443-458. DOI:10.3969/j.issn.1674-3466.2006.05.002 |
王艳红, 王珂, 邢福. 2005. 匍匐茎草本植物形态可塑性、整合作用与觅食行为研究进展. 生态学杂志, 24(1): 70-74. |
温达志, 孔国辉, 林植芳, 等. 1999. 光强对四种亚热带树苗生长特征的影响的比较. 热带亚热带植物学报, 7(2): 125-132. |
肖春旺, 刘玉成. 1999. 不同光环境的四川大头茶幼苗的生态适应. 生态学报, 19(3): 422-426. DOI:10.3321/j.issn:1000-0933.1999.03.022 |
徐程扬. 2001. 不同光环境下紫椴幼树树冠结构的可塑性响应. 应用生态学报, 12(3): 339-343. DOI:10.3321/j.issn:1001-9332.2001.03.005 |
Agyeman V K, Swaine M D, Thompson J. 1999. Response of tropical forest tree seedlings to irradiance and the derivation of a light response index. Journal of Ecology, 87: 815-827. DOI:10.1046/j.1365-2745.1999.00400.x |
Canham C D. 1998. Growth and canopy architecture of shade-toLerant trees:response to canopy gaps. Ecology, 69(3): 786-795. |
Gardiner E S, Hodges J D. 1998. Growth and biomass distribution of cherrybark oak (Quercus pagoda Raf.) seedlings as influenced by light availability. For Ecological Man, 108: 127-134. |
Gilmore D W, Seymour R S. 1997. Crown architecture of Abies balsamea from four canopy positions. Tree Physiology, 17: 71-80. DOI:10.1093/treephys/17.2.71 |
Hart J W. 1998. Light and plant Growth. London: Unwin Hyman, 1-204.
|
King D A. 2003. Allocation of above-ground growth is related to light in temperate deciduous saplings. Functional Ecology, 17(4): 482-488. DOI:10.1046/j.1365-2435.2003.00759.x |
Pearcy R W. 1999. Responses of plants to heterogeneous light environments//Pugnaire F I, Vallsdares F(eds). Handbook of Functional Plant Ecology. New York: Marcel Dekker Inc, 270-314.
|
Peters R. 1997. Beech forests. Dordrecht: Kluwer Academic Publishers.
|
Poorter L. 1999. Growth response of 15 rain-forest tree species to a light gradient: The relative importance of morphological and physiological traits. Function al Ecology, 13: 396-410. DOI:10.1046/j.1365-2435.1999.00332.x |
Rosenthal J P, Kotane P M. 1994. Terrestrial plant tolerance to herbivory. Trends in Ecology and Evolution, 9(4): 145-148. DOI:10.1016/0169-5347(94)90180-5 |
Sack L, Grubb P J. 2002. The combined impacts of deep shade and drought on the growth and biomass allocation of shade-tolerant woody seedings. Oecologia, 131: 175-185. DOI:10.1007/s00442-002-0873-0 |